Caracteristicile structurii țesutului conjunctiv. Țesutul conjunctiv (anatomia umană). Structura țesutului conjunctiv
Țesut conjunctiv format în timpul dezvoltării embrionare din mezenchim. Îndeplinește o serie de funcții importante în organism: - Suport-mecanic- datorita faptului ca tesutul conjunctiv este cadrul de sustinere a intregului corp (schelet) si a majoritatii organelor;
- Trofic (metabolic)- este determinată de faptul că țesutul conjunctiv însoțește peste tot vasele de sânge (chiar și pe cele mai mici) și este un intermediar în implementarea proceselor metabolice dintre sânge și alte țesuturi ale organelor.
- De protecţie- depinde de prezența fagocitelor în țesutul conjunctiv, care participă activ la reacțiile imune.
- Reparator (plastic)- se manifestă prin participarea activă a țesutului conjunctiv la procesele de regenerare (refacerea integrității țesuturilor și organelor după lezare sau boală).
Țesutul conjunctiv reprezintă mai mult de jumătate din greutatea corporală, iar gradul de dezvoltare a acestuia crește treptat în timpul evoluției vertebratelor. Potrivit academicianului A. A. Bogomolets; țesutul conjunctiv formează un singur sistem, care are o distribuție uriașă și se caracterizează prin reproducerea și migrarea rapidă a celulelor către partea dorită a corpului, interacțiunea lor, participarea activă la fenomenele de regenerare și reacții imune.
O particularitate a structurii țesutului conjunctiv este prezența în acesta, împreună cu celulele, a unei substanțe intercelulare foarte dezvoltate (substanță amorfă și fibre). Pe baza structurii substanței intercelulare, se pot distinge principalele tipuri de țesut conjunctiv:
Sânge și limfa
Nu există structuri fibroase în substanța intercelulară a sângelui (plasmă) și limfei, prin urmare aceste tipuri de țesut conjunctiv au o consistență lichidă. Compoziția chimică a limfei este apropiată de plasmă, care conține așa-numitele elemente formate: globule roșii (eritrocite), globule albe (leucocite) și trombocite (trombocite). La mamifere, dintre elementele de formă enumerate, doar leucocitele sunt la fel. Globulele roșii sunt reprezentate de formațiuni post-celulare, deoarece, dezvoltându-se din celule care au nuclei, le pierd în procesul de acumulare a hemoglobinei. Trombocitele la mamifere sunt fragmente de celule gigantice speciale (megacariocite), care se găsesc în principal în măduva osoasă roșie. Leucocitele sunt împărțite în granulare (granulocite) și negranulare (agranulocite). La rândul lor, granulocitele sunt legate de baze acide sau coloranți neutri, deci sunt împărțite în oxifile, bazofile și neutrofile. Agranulocitele se diferențiază în limfocite și monocite.Funcția principală a sângelui este transportul, ale cărui manifestări sunt trofismul, respirația, protecția și homeostazia.
Țesut conjunctiv reticular
Țesutul conjunctiv reticular (din latinescul Reticulum) are o structură de plasă, deoarece este format din celule care au o formă de găleată și sunt în contact unele cu altele cu procesele lor.Printre celulele sale se numără fibroblastele, care produc substanță intercelulară, și macrofagele fixe, care se formează din monocite din sânge. Substanța intercelulară a țesutului reticular este reprezentată de o substanță amorfă și de fibre reticulare subțiri, care sunt un tip de colagen. Țesutul reticular face parte din organele hematopoietice și are o consistență semi-lichidă. Participă la hematopoieză, creând un mediu (micromediu) pentru celulele sanguine care se dezvoltă și îndeplinește o funcție de protecție cu ajutorul fagocitelor.
Țesut conjunctiv fibros
În țesutul conjunctiv fibros, numărul de fibre poate fi moderat (țesut fibros lax) sau mai semnificativ (țesut fibros dens). Țesutul conjunctiv fibros lax este cel mai comun tip de țesut conjunctiv. Îndeplinește predominant funcții trofice și de protecție și participă la aproape toate reacțiile fiziologice și de protecție care apar în organism. Celulele țesutului conjunctiv lax includ fibroblaste, macrofagocite, plasmocite (plasmocite), bazofile tisulare, adipocite (celule adipoase) și pigmentocite (celule pigmentare).Cel mai numeros grup de celule ale țesutului conjunctiv sunt fibroblastele, care formează substanța intermediară (substanța și fibrele amorfe). Fibroblastele au o ultrastructură specială, tipică celulelor secretoare (conțin un reticul endoplasmatic granular bine dezvoltat și complex Golgi). Fibroblastii tineri sunt capabili să se divizeze și au o formă de găleată. Fibroblastele mature (fibrocitele) nu se pot diviza. Majoritatea organelelor lor sunt reduse, iar activitatea lor funcțională este semnificativ redusă.
Al doilea grup ca mărime de celule de țesut conjunctiv este macrofagocitele. Ele sunt formate din monocite din sânge, în care, în procesul de transformare în macrofage tisulare, crește conținutul de organite, precum și lizozomi. Aceste modificări reflectă capacitatea macrofagelor tisulare pentru fagocitoză energetică și sinteza unui număr de substanțe biologic active.
Celule plasmatice sunt formate din limfocitele B. Au o formă rotundă sau ovală, un nucleu situat excentric cu pete de heterocromatină orientate radial, un reticul endoplasmatic granular bine dezvoltat și un complex Golgi, situat în apropierea nucleului (în zona citoplasmei care a pierdut reticulul granular) . Funcția acestor celule este sinteza anticorpilor - o proteină specială (gamma globulină) care neutralizează antigenele din organism.
Bazofilele tisulare conțin granule bazofile în citoplasmă. Aceste celule secretă așa-numitele amine biogene (heparină, histamina, serotonina), care sunt implicate în reglarea coagulării sângelui, a permeabilității tisulare a vaselor mici de sânge etc. În acest sens, bazofilele tisulare sunt regulatori ai homeostaziei locale.
Adipocite(celulele adipoase) sunt capabile să depună grăsime, care este o substanță energetică și trofică, un izolator termic. Există adipocite de grăsime albă și maro. Adipocitele de grăsime albă au o incluziune mare de grăsime (picătură) formată din grăsime neutră, unde cea mai mare parte a citoplasmei cu nucleul este împinsă către unul dintre polii celulei.
Adipocitele de grăsime brună au dimensiuni mai mici. Nucleul din ele este situat în centru, iar grăsimea se află în citoplasmă sub formă de numeroase picături mici. Celulele adipoase brune se caracterizează prin faptul că conțin un număr mare de mitocondrii. Funcția principală a țesutului adipos brun este de a genera căldură. La om, este bine dezvoltat în primele luni de viață, iar apoi este înlocuit cu țesut adipos alb.
Pigmentocite(celule pigmentare) - elemente celulare în formă de găleată care conțin granule de pigment - melanina - în citoplasmă. La oameni și mamifere, aceștia fac parte din piele (atât dermul, cât și epiderma), foliculii de păr, meningele, irisul și coroida. Aceste celule îndeplinesc o funcție de protecție, reducând efectele nocive ale razelor solare asupra pielii și organului de vedere.
Țesutul este o colecție de celule și substanțe intercelulare care au aceeași structură, funcție și origine.
În corpul mamiferelor, animalelor și omului, există 4 tipuri de țesuturi: epitelial, conjunctiv, în care se pot distinge osul, cartilajul și țesutul adipos; musculoși și nervoși.
Țesut - localizare în organism, tipuri, funcții, structură
Țesuturile sunt un sistem de celule și substanțe intercelulare care au aceeași structură, origine și funcții.
Substanța intercelulară este un produs al activității celulare. Acesta asigură comunicarea între celule și creează un mediu favorabil pentru acestea. Poate fi lichid, cum ar fi plasma sanguină; amorf - cartilaj; structurat - fibre musculare; țesut osos dur (sub formă de sare).
Celulele tisulare au forme diferite, care le determină funcția. Țesăturile sunt împărțite în patru tipuri:
- țesuturi epiteliale - de frontieră: piele, mucoasă;
- conjunctiv - mediul intern al corpului nostru;
- muşchi;
- țesut nervos.
Tesut epitelial
Țesuturile epiteliale (de frontieră) - acoperă suprafața corpului, membranele mucoase ale tuturor organelor și cavitățile interne ale corpului, membranele seroase și formează, de asemenea, glandele secreției externe și interne. Epiteliul care căptușește membrana mucoasă este situat pe membrana bazală, iar suprafața sa interioară se confruntă direct cu mediul extern. Nutriția sa se realizează prin difuzarea substanțelor și a oxigenului din vasele de sânge prin membrana bazală.
Caracteristici: sunt multe celule, există puțină substanță intercelulară și este reprezentată de o membrană bazală.
Țesuturile epiteliale îndeplinesc următoarele funcții:
- de protecţie;
- excretor;
- aspiraţie
Clasificarea epiteliilor. Pe baza numărului de straturi, se face o distincție între un singur strat și mai multe straturi. Se clasifică după formă: plate, cubice, cilindrice.
Dacă toate celulele epiteliale ajung la membrana bazală, este un epiteliu cu un singur strat, iar dacă doar celulele dintr-un rând sunt conectate la membrana bazală, în timp ce altele sunt libere, este multistrat. Epiteliul cu un singur strat poate fi cu un singur rând sau cu mai multe rânduri, ceea ce depinde de nivelul de localizare a nucleelor. Uneori, epiteliul mononuclear sau multinuclear are cili ciliați îndreptați spre mediul extern.
Epiteliu stratificat Țesutul epitelial (tegumentar), sau epiteliul, este un strat limită de celule care acoperă tegumentul corpului, membranele mucoase ale tuturor organelor și cavităților interne și formează, de asemenea, baza multor glande.
Epiteliul glandular Epiteliul separă organismul (mediul intern) de mediul extern, dar în același timp servește ca intermediar în interacțiunea organismului cu mediul. Celulele epiteliale sunt strâns legate între ele și formează o barieră mecanică care împiedică pătrunderea microorganismelor și a substanțelor străine în organism. Celulele țesutului epitelial trăiesc pentru o perioadă scurtă de timp și sunt rapid înlocuite cu altele noi (acest proces se numește regenerare).
Țesutul epitelial este implicat și în multe alte funcții: secreție (glande exocrine și endocrine), absorbție (epiteliul intestinal), schimbul de gaze (epiteliul pulmonar).
Caracteristica principală a epiteliului este că constă dintr-un strat continuu de celule strâns adiacente. Epiteliul poate fi sub forma unui strat de celule care căptușește toate suprafețele corpului și sub formă de acumulari mari de celule - glande: ficat, pancreas, tiroida, glande salivare etc. În primul caz, se află pe membrana bazală, care separă epiteliul de țesutul conjunctiv subiacent. Cu toate acestea, există și excepții: celulele epiteliale din țesutul limfatic alternează cu elemente de țesut conjunctiv; un astfel de epiteliu este numit atipic.
Celulele epiteliale, dispuse într-un strat, se pot afla în mai multe straturi (epiteliu stratificat) sau într-un singur strat (epiteliu cu un singur strat). Pe baza înălțimii celulelor, epiteliile sunt împărțite în plate, cubice, prismatice și cilindrice.
Epiteliu scuamos cu un singur strat - căptușește suprafața membranelor seroase: pleura, plămânii, peritoneul, pericardul inimii.
Epiteliu cubic cu un singur strat - formează pereții tubilor renali și canalele excretoare ale glandelor.
Epiteliul columnar cu un singur strat - formează mucoasa gastrică.
Epiteliu mărginit - un epiteliu cilindric cu un singur strat, pe suprafața exterioară a celulelor a cărui margine este formată din microvilozități care asigură absorbția nutrienților - căptușește membrana mucoasă a intestinului subțire.
Epiteliul ciliat (epiteliul ciliat) este un epiteliu pseudostratificat format din celule cilindrice, a cărui margine interioară, adică îndreptată spre cavitate sau canal, este echipată cu formațiuni asemănătoare părului (cili) care oscilează constant - cilii asigură mișcarea oului în tuburile; elimină germenii și praful din tractul respirator.
Epiteliul stratificat este situat la granița dintre organism și mediul extern. Dacă în epiteliu apar procese de keratinizare, adică straturile superioare ale celulelor se transformă în solzi cornos, atunci un astfel de epiteliu multistrat se numește keratinizare (suprafața pielii). Epiteliul multistrat căptușește membrana mucoasă a gurii, cavitatea alimentară și corneea ochiului.
Epiteliul de tranziție căptușește pereții vezicii urinare, pelvisului renal și ureterului. Când aceste organe sunt umplute, epiteliul de tranziție se întinde și celulele se pot muta de la un rând la altul.
Epiteliul glandular – formează glande și îndeplinește o funcție secretorie (eliberează substanțe – secreții care fie sunt eliberate în mediul extern, fie pătrund în sânge și limfă (hormoni)). Capacitatea celulelor de a produce si secreta substante necesare functionarii organismului se numeste secretie. În acest sens, un astfel de epiteliu a fost numit și epiteliu secretor.
Țesut conjunctiv
Țesutul conjunctiv Constă din celule, substanță intercelulară și fibre de țesut conjunctiv. Este format din oase, cartilaje, tendoane, ligamente, sânge, grăsime, este prezent în toate organele (țesut conjunctiv lax) sub forma așa-numitei strome (cadru) de organe.
Spre deosebire de țesutul epitelial, în toate tipurile de țesut conjunctiv (cu excepția țesutului adipos), substanța intercelulară predomină asupra celulelor în volum, adică substanța intercelulară este foarte bine exprimată. Compoziția chimică și proprietățile fizice ale substanței intercelulare sunt foarte diverse în diferite tipuri de țesut conjunctiv. De exemplu, sângele - celulele din el „plutesc” și se mișcă liber, deoarece substanța intercelulară este bine dezvoltată.
În general, țesutul conjunctiv formează ceea ce se numește mediul intern al corpului. Este foarte divers și este reprezentat de diferite tipuri - de la forme dense și libere până la sânge și limfă, ale căror celule se află în lichid. Diferențele fundamentale între tipurile de țesut conjunctiv sunt determinate de raporturile componentelor celulare și de natura substanței intercelulare.
Țesutul conjunctiv fibros dens (tendoane musculare, ligamente articulare) este dominat de structuri fibroase și suferă un stres mecanic semnificativ.
Țesutul conjunctiv fibros lax este extrem de comun în organism. Este foarte bogat, dimpotrivă, în forme celulare de diferite tipuri. Unele dintre ele sunt implicate în formarea fibrelor tisulare (fibroblaste), altele, ceea ce este deosebit de important, asigură în primul rând procese de protecție și reglare, inclusiv prin mecanisme imunitare (macrofage, limfocite, bazofile tisulare, plasmocite).
Os
Țesutul osos Țesutul osos, care formează oasele scheletului, este foarte durabil. Menține forma corpului (constituția) și protejează organele situate în craniu, torace și cavitățile pelvine și participă la metabolismul mineral. Țesutul este format din celule (osteocite) și substanță intercelulară în care se află canalele nutritive cu vase de sânge. Substanța intercelulară conține până la 70% săruri minerale (calciu, fosfor și magneziu).
În dezvoltarea sa, țesutul osos trece prin stadii fibroase și lamelare. În diferite părți ale osului este organizat sub formă de substanță osoasă compactă sau spongioasă.
Țesutul cartilajului
Țesutul cartilajului este format din celule (condrocite) și substanță intercelulară (matricea cartilajului), caracterizată prin elasticitate crescută. Îndeplinește o funcție de susținere, deoarece formează cea mai mare parte a cartilajului.
Există trei tipuri de țesut cartilaginos: hialin, care face parte din cartilajul traheei, bronhiile, capetele coastelor și suprafețele articulare ale oaselor; elastic, formând auricul și epiglota; fibros, situat în discurile intervertebrale și articulațiile oaselor pubiene.
Țesut adipos
Țesutul adipos este similar cu țesutul conjunctiv lax. Celulele sunt mari și pline de grăsime. Țesutul adipos îndeplinește funcții nutriționale, de formare a formei și de termoreglare. Țesutul adipos este împărțit în două tipuri: alb și maro. La om predomină țesutul adipos alb, o parte din acesta înconjoară organele, menținându-și poziția în corpul uman și alte funcții. Cantitatea de țesut adipos maro la om este mică (se găsește în principal la nou-născuți). Funcția principală a țesutului adipos brun este producerea de căldură. Țesutul adipos maro menține temperatura corpului animalelor în timpul hibernării și temperatura nou-născuților.
Muşchi
Celulele musculare sunt numite fibre musculare deoarece sunt întinse în mod constant într-o direcție.
Clasificarea țesutului muscular se realizează pe baza structurii țesutului (histologic): prin prezența sau absența striațiilor transversale și pe baza mecanismului de contracție - voluntar (ca în mușchiul scheletic) sau involuntar (neted). sau mușchiul cardiac).
Țesutul muscular are excitabilitate și capacitatea de a se contracta activ sub influența sistemului nervos și a anumitor substanțe. Diferențele microscopice ne permit să distingem două tipuri de acest țesut - neted (nestriat) și striat (striat).
Țesutul muscular neted are o structură celulară. Formează membranele musculare ale pereților organelor interne (intestine, uter, vezică urinară etc.), vaselor sanguine și limfatice; contractia lui are loc involuntar.
Țesutul muscular striat este format din fibre musculare, fiecare dintre acestea fiind reprezentată de multe mii de celule, fuzionate, pe lângă nucleele lor, într-o singură structură. Formează mușchii scheletici. Le putem scurta după bunul plac.
Un tip de țesut muscular striat este mușchiul cardiac, care are abilități unice. În timpul vieții (aproximativ 70 de ani), mușchiul inimii se contractă de peste 2,5 milioane de ori. Nicio altă țesătură nu are un asemenea potențial de rezistență. Țesutul muscular cardiac are striații transversale. Cu toate acestea, spre deosebire de mușchiul scheletic, există zone speciale unde fibrele musculare se întâlnesc. Datorită acestei structuri, contracția unei fibre se transmite rapid celor vecine. Acest lucru asigură contracția simultană a unor zone mari ale mușchiului inimii.
De asemenea, caracteristicile structurale ale țesutului muscular sunt că celulele sale conțin mănunchiuri de miofibrile formate din două proteine - actină și miozină.
tesut nervos
Țesutul nervos este format din două tipuri de celule: nervoase (neuroni) și gliale. Celulele gliale sunt strâns adiacente neuronului, îndeplinind funcții de susținere, nutriție, secretoare și de protecție.
Neuronul este unitatea structurală și funcțională de bază a țesutului nervos. Caracteristica sa principală este capacitatea de a genera impulsuri nervoase și de a transmite excitația altor neuroni sau celulelor musculare și glandulare ale organelor de lucru. Neuronii pot consta dintr-un corp și procese. Celulele nervoase sunt concepute pentru a conduce impulsurile nervoase. După ce a primit informații pe o parte a suprafeței, neuronul o transmite foarte rapid către o altă parte a suprafeței sale. Deoarece procesele unui neuron sunt foarte lungi, informațiile sunt transmise pe distanțe mari. Majoritatea neuronilor au două tipuri de procese: scurte, groase, ramificate în apropierea corpului - dendrite și lungi (până la 1,5 m), subțiri și ramificați doar la capăt - axonii. Axonii formează fibre nervoase.
Un impuls nervos este o undă electrică care se deplasează cu viteză mare de-a lungul unei fibre nervoase.
În funcție de funcțiile îndeplinite și de caracteristicile structurale, toate celulele nervoase sunt împărțite în trei tipuri: senzoriale, motorii (executive) și intercalare. Fibrele motorii care circulă ca parte a nervilor transmit semnale către mușchi și glande, fibrele senzoriale transmit informații despre starea organelor către sistemul nervos central.
Acum putem combina toate informațiile primite într-un tabel.
Tipuri de țesături (tabel)
|
Grup de țesături |
Tipuri de țesături |
Structura tisulară |
Locație |
|
| Epiteliu | Apartament | Suprafața celulelor este netedă. Celulele sunt strâns adiacente între ele | Suprafața pielii, cavitatea bucală, esofag, alveole, capsule de nefron | Tegumentar, protector, excretor (schimb gazos, excreție urinară) |
| Glandular | Celulele glandulare produc secreții | Glande cutanate, stomac, intestine, glande endocrine, glande salivare | Excretor (secreție de transpirație, lacrimi), secretorie (formare de salivă, suc gastric și intestinal, hormoni) | |
| ciliat (ciliat) | Constă din celule cu numeroase fire de păr (cili) | Căile aeriene | Protectie (cilii capteaza si indeparteaza particulele de praf) | |
| Conjunctiv | Fibroasă densă | Grupuri de celule fibroase, strâns împachetate, fără substanță intercelulară | Pielea în sine, tendoanele, ligamentele, membranele vaselor de sânge, corneea ochiului | Tegumentar, protector, motor |
| Fibroase libere | Celule fibroase aranjate lejer, împletite între ele. Substanța intercelulară este lipsită de structură | Țesut adipos subcutanat, sac pericardic, căi ale sistemului nervos | Conectează pielea de mușchi, susține organele din corp, umple golurile dintre organe. Oferă termoreglarea corpului | |
| Cartilaginos | Celule vii rotunde sau ovale situate în capsule, substanța intercelulară este densă, elastică, transparentă | Discuri intervertebrale, cartilaj laringian, trahee, auricul, suprafața articulară | Netezirea suprafețelor de frecare ale oaselor. Protecție împotriva deformării căilor respiratorii și a urechilor | |
| Os | Celule vii cu procese lungi, interconectate, substanță intercelulară - săruri anorganice și proteină oseină | Oasele scheletului | Suport, motor, protector | |
| Sânge și limfa | Țesutul conjunctiv lichid este format din elemente formate (celule) și plasmă (lichid cu substanțe organice și minerale dizolvate în el - ser și proteină fibrinogen) | Sistemul circulator al întregului organism | Transportă O2 și nutrienți în tot corpul. Colectează CO 2 și produse de disimilare. Asigură constanța mediului intern, compoziția chimică și gazoasă a organismului. Protectoare (imunitate). Regulator (umoral) | |
| Muscular | Cu dungi încrucișate | Celule cilindrice multinucleate de până la 10 cm lungime, striate cu dungi transversale | Mușchii scheletici, mușchii cardiaci | Mișcări voluntare ale corpului și părților sale, expresii faciale, vorbire. Contracții involuntare (automate) ale mușchiului inimii pentru a împinge sângele prin camerele inimii. Are proprietăți de excitabilitate și contractilitate |
| Neted | Celule mononucleare de până la 0,5 mm lungime cu capete ascuțite | Pereții tractului digestiv, vasele de sânge și limfatice, mușchii pielii | Contracții involuntare ale pereților organelor interne goale. Ridicarea părului pe piele | |
| agitat | Celulele nervoase (neuroni) | Corpi de celule nervoase, variate ca formă și dimensiune, până la 0,1 mm în diametru | Formează substanța cenușie a creierului și a măduvei spinării | Activitate nervoasă mai mare. Legătura corpului cu Mediul extern. Centre de reflexe condiționate și necondiționate. Țesutul nervos are proprietăți de excitabilitate și conductivitate |
| Procese scurte ale neuronilor - dendrite ramificate | Conectați-vă cu procesele celulelor învecinate | Ele transmit excitația unui neuron altuia, stabilind o legătură între toate organele corpului | ||
| Fibre nervoase - axoni (neurite) - procese lungi de neuroni de până la 1,5 m lungime. Organele se termină cu terminații nervoase ramificate | Nervi ai sistemului nervos periferic care inervează toate organele corpului | Căile sistemului nervos. Ei transmit excitația de la celula nervoasă la periferie prin intermediul neuronilor centrifugi; de la receptori (organe inervate) - până la celula nervoasă de-a lungul neuronilor centripeți. Interneuronii transmit excitația de la neuronii centripeți (sensibili) la neuronii centrifugi (motorii) |
1) Funcția trofică.
Țesutul conjunctiv superficial acoperă toate vasele de sânge, astfel încât schimbul de substanțe între sânge și orice alt țesut are loc cu participarea obligatorie a țesutului conjunctiv. În esență, țesutul conjunctiv lax reglează schimbul de substanțe între sânge și alte țesuturi.
2) Funcția de suport.
Există două tipuri de funcție de sprijin: stromală și formatoare.
- Funcția stromală.
Țesutul conjunctiv lax formează stroma - cadrul organelor interne.
- Formarea formei.
Țesutul conjunctiv dens formează capsula organului, care formează forma organului.
3) Funcție de protecție.
Țesutul conjunctiv îndeplinește mai degrabă funcțiile de protecție imunitară decât de protecție mecanică. Apărarea imună este realizată de macrofage, mastocite și anticorpi produși de țesutul conjunctiv. Deși țesutul conjunctiv dens poate servi drept protecție mecanică.
4) Funcția mecanică este funcția de organizare a mișcării.
Țesutul conjunctiv dens formează tendoane și ligamente care participă la organizarea sistemului musculo-scheletic.
5) Funcție plastică.
Participarea țesutului conjunctiv lax la organizarea regenerării. Dacă în timpul procesului de activitate vitală se formează vreun defect sau răni într-un organ, acestea sunt umplute cu țesut conjunctiv lax. Ca urmare, forma organului este restabilită.
Reglarea funcțiilor țesutului conjunctiv se realizează la toate nivelurile de organizare - la nivelul unei celule, al unui organ, al unui organism. La nivel celular, contactele intercelulare sunt importante printr-o substanță efectoră strâns asociată cu membrana celulară, și mediatori eliberați în spațiul intercelular: limfokine, monokine, fibrokine, labrokine (respectiv, mediatori ai limfocitelor, monocitelor, fibroblastelor, bazofilelor tisulare). Pe lângă mediatorii specifici, pentru care există receptori corespunzători pe membranele celulare, există și mediatori nespecifici - prostaglandine, muramidaze, fibronectină, proteaze.
Relația dintre elementele țesutului conjunctiv se realizează conform principiului feedback-ului, care în condiții normale asigură adecvarea răspunsurilor, iar în patologie, adaptabilitate și fiabilitate ridicate. Autoreglementarea „de jos”, bazată pe interacțiuni cooperante între celule, este completată de reglarea endocrină și nervoasă, construită după principiul ierarhic „de sus în jos”.
În acest sens, un rol important îi revine hormonului hipofizei anterioare - somatotropina. Stimulează reproducerea celulelor țesutului conjunctiv și procesele sintetice din ele. În același timp, corticotropina și glicocorticoizii inhibă proliferarea, provoacă diferențierea și maturarea prematură a fibroblastelor, care este însoțită de colagenogeneză afectată. Rolul insulinei în reglarea structurii și funcției țesutului conjunctiv este că accelerează schimbul de acid hialuronic și sulfat de condroitin. Aparent, acest lucru explică încălcările grave ale țesutului conjunctiv în general și ale peretelui vascular, în special, în diabetul zaharat (angiopatie diabetică).
Dereglarea sistemului de țesut conjunctiv poate apărea și la orice nivel. În țesutul conjunctiv, relațiile de autoreglare stabilite și conexiunile sale cu celulele parenchimului pot fi perturbate. Tulburarea se poate manifesta predominant într-unul sau altul organ (articulații în reumatism, piele în lupusul eritematos sistemic). În fine, dereglarea țesutului conjunctiv se poate manifesta în toate organele și în organism în ansamblu (proliferarea osoasă în acromegalie, nanism și edem mucos în hipotiroidie).
Starea țesutului conjunctiv joacă un rol important în timpul îmbătrânirii. La un moment dat, A. A. Bogomolets a remarcat că „îmbătrânirea începe tocmai cu țesutul conjunctiv”. El a văzut motivul îmbătrânirii în faptul că apar modificări fizice și chimice ale componentelor macromoleculare - „maturarea coloizilor celulari și a miceloizilor, transformarea lor în precipitate și floculate, formând incluziuni inerte biologic care inhibă activitatea vitală a celulelor”.
Cercetările moderne au arătat că odată cu vârsta, apar modificări în țesutul conjunctiv care îi perturbă funcțiile trofice, de protecție și alte funcții. În piele, tendoane, țesutul cartilajului și aortă, numărul de celule, dimensiunea lor și dimensiunea nucleelor scade. Reproducerea fibroblastelor, de exemplu, în timpul inflamației la persoanele în vârstă este încetinită. În structurile fibroase, modificările legate de vârstă sunt exprimate printr-o creștere a numărului de legături încrucișate între fibrele de colagen. Colagenul cu o cantitate în exces de „legături” macromoleculare capătă noi proprietăți. Devine mai rezistent la influențele temperaturii, solubilitatea și capacitatea de a lega apa scade. În mod normal, stabil din punct de vedere metabolic, colagenul devine din ce în ce mai inert. Auto-reînnoirea sa încetinește, ceea ce duce inevitabil la acumularea de noi erori în molecule. Cantitatea de acid hialuronic din substanța principală scade, ceea ce probabil reduce capacitatea organismului de a lega apa odată cu vârsta. În același timp, cantitatea de sulfat de condroitină din peretele vascular crește. Acesta din urmă favorizează calcificarea vasculară, deoarece glicozaminoglicanii sulfatați au o afinitate pentru ionii de calciu.
Funcția de protecție a țesutului conjunctiv este exprimată prin faptul că, prin participarea la structura pielii, a membranei mucoase, a capsulelor fibroase, precum și la structurile barierelor specializate (glia din bariera hemato-encefalică), contribuie la crearea de bariere mecanice. Celulele țesutului conjunctiv determină o astfel de formă de protecție precum fagocitoza (macrofagocite), precum și capacitatea de a delimita țesutul deteriorat de țesutul normal (arborele de granulație). În cele din urmă, celulele țesutului conjunctiv, care interacționează cu limfocitele, participă la răspunsul imun.
A. A. Bogomolets a fost primul care a apreciat pe deplin rolul protector al țesutului conjunctiv. El și studenții săi au arătat că reactivitatea scăzută a țesutului conjunctiv, de regulă, este combinată cu un curs mai sever al bolii: rănile se vindecă mai lent, fracturile se vindecă mai puțin bine.
Funcțiile țesutului conjunctiv
Toate acestea ne-au permis să presupunem că prin influențarea țesutului conjunctiv este posibilă agravarea sau îmbunătățirea cursului bolii, făcând-o mai puțin gravă. A fost nevoie doar de a găsi o modalitate de a-i stimula funcțiile. Anumite rezultate puteau fi obținute prin metode precum kinetoterapie, masaj, insolație, dietă, dar A. A. Bogomolets a avut în vedere stimularea selectivă specifică a tuturor elementelor țesutului conjunctiv, oriunde se află acestea în organism. Ca urmare, acest lucru a fost realizat imunologic, adică. injectarea la oameni a serului obținut ca urmare a imunizării animalelor (cailor) cu organe bogate în țesut conjunctiv (măduvă osoasă, splină). Acest ser a primit denumirea prescurtată ACS (ser antireticular citotoxic) și a fost studiat în detaliu de A. A. Bogomolets și studenții săi, iar apoi folosit în clinică.
S-a constatat că efectul serului citotoxic antireticular depinde de doza acestuia. Doze mari de ser au avut un efect citotoxic, adică. distruge celulele țesutului conjunctiv, mici - stimulatoare. Acest lucru a fost exprimat printr-o creștere a metabolismului în elementele macrofagocitare ale țesutului și activarea fagocitozei, o creștere a titrului de anticorpi antimicrobieni și normalizarea metabolismului apă-electroliți și grăsimi. Activarea proceselor enzimatice permite țesutului conjunctiv să se elibereze de substanțele de balast care se acumulează în el în timpul diferitelor boli metabolice, precum și în timpul îmbătrânirii. În acest caz se pot forma substanțe care au un efect nespecific asupra celulelor organismului, constituind un efect general de stimulare.
Caută Prelegeri
Funcțiile mecanice ale scheletului
1. Funcția de suport constă în faptul că oasele susțin țesuturile moi atașate de ele (mușchi, fascia și alte organe) și participă la formarea pereților cavităților în care se află organele interne.
2. Funcția arc datorită prezenței în schelet a unor formațiuni care atenuează șocurile și șocurile (tampoane cartilaginoase, cartilaj articular între oasele de legătură etc.)
3. Funcția de protecție este că scheletul formează recipiente pentru organele vitale și le protejează de influențele externe.
4. Funcția motorului posibil datorită structurii oaselor sub formă de pârghii lungi și scurte conectate prin articulații mobile și antrenate de mușchi controlați de sistemul nervos.
5. Funcție anti-gravitație se manifestă prin faptul că scheletul creează suport pentru stabilitatea corpului care se ridică deasupra solului.
În plus, oasele determină direcția cursului vaselor de sânge, nervilor și mușchilor, precum și forma corpului și dimensiunea acestuia.
Funcțiile biologice ale scheletului
1. Funcții metabolice- scheletul este implicat in metabolism (mai ales in metabolismul mineral), fiind un depozit de saruri minerale - fosfor, calciu, fier etc.
2. Funcția hematopoietică datorită faptului că în interiorul oaselor se află măduva osoasă roșie - organul hematopoietic central - partea organică a osului.
3. Funcția imunologică este, de asemenea, asociat cu măduva osoasă roșie: aceasta din urmă conține o populație auto-susținută de celule stem hematopoietice, din care se formează și celule ale sistemului imunitar sau limfocite.
Osul ca organ
Fiecare os tubular are următoarele părți:
1. Diafiza(corp osos) este un tub osos care conține măduvă osoasă galbenă la adulți și îndeplinește funcția de sprijin și respectiv de protecție.
2. Metafize(capete ale diafizei), adiacente cartilajului metaepifizar, se dezvoltă împreună cu diafiza, dar participă la creșterea oaselor în lungime și constau din substanță spongioasă.
3. Epifize(capete articulare ale fiecărui os tubular) sunt situate pe cealaltă parte a cartilajului metaepifizar.
4. Apofize(proeminențe osoase situate în apropierea epifizei).
Clasificarea oaselor
Numărul de oase individuale care alcătuiesc scheletul uman adult este mai mare de 200 (206 oase). Oasele variază în mărime și formă și ocupă poziții specifice în corp. Pe baza formei lor externe, oasele se disting între lungi, scurte, late și mixte.
Cu toate acestea, este mai corect să distingem oasele pe baza a trei principii pe care se bazează orice clasificare anatomică - formă (structură), funcție și dezvoltare. Din acest punct de vedere, se disting următoarele grupe de oase:
![]()
![]() OASE
OASE
Tubular spongios plat mixt aeropurtat
Oasele craniului lung lungi scurte
Centuri osoase sesamoide scurte
Conexiunea oaselor
Există trei tipuri de conexiuni osoase:
1) Conexiuni continue (sinartroză), când există un strat de țesut conjunctiv sau cartilaj între oase. Nu există nici un spațiu sau cavitate între oasele de legătură.
2) Conexiuni intermitente sau articulații (diartroză sau articulații sinoviale) - când există o cavitate între oase și o membrană sinovială care căptușește interiorul capsulei articulare.
3) Semi-articulații sau simfize (hemiartroză), când există un mic decalaj în stratul cartilaginos sau de țesut conjunctiv între oasele de legătură.
1. Legături continue – sinartroză. În funcție de structura țesutului care leagă oasele, se disting următoarele grupuri de aceste conexiuni:
— țesut fibros (sindesmoze) sau conjunctiv;
— cartilaginoase (sincondroză);
- conexiuni osoase (sinostoze);
- elastic;
- conexiuni musculare.
Articulații fibroase (sindesmoze) Acestea sunt conexiuni puternice prin țesutul conjunctiv fibros dens. Acestea includ:
A) membrane sau membrane interoase.
b) mănunchiuri
V) cusaturi:
- zimțat (de exemplu, legătura dintre oasele frontale și parietale);
- solzoase (de exemplu, legătura osului temporal cu parietalul);
- netede (de exemplu, conexiuni între oasele craniului facial)/
G) ciocănind
Articulațiile cartilajului (sincondroză) Sunt conexiuni între oase și cartilaj. În funcție de durata existenței lor, sincondroza este:
A) temporar– există până la o anumită vârstă, după care sunt înlocuite de sinostoze (de exemplu, între oasele centurii pelvine).
b) permanent– există până la o anumită vârstă, după care sunt înlocuite cu sinostoze (de exemplu, între piramida osului temporal și oasele adiacente ale centurii pelvine);
Conexiuni elastice nu au rezistența pe care o au țesutul conjunctiv sau compușii fibroși.
Articulații osoase (sinostoze): în spațiul dintre oase, țesutul conjunctiv se transformă în os sau mai întâi în cartilaj și apoi în os.
Conexiuni musculare Ele sunt mobile și variabile în lungime conexiuni a două sau mai multe oase cu ajutorul mușchilor striați.
2. Articulații discontinue sau articulații (diartroză) sunt cele mai avansate tipuri de conexiuni osoase.
În fiecare articulație se disting următoarele: elemente esentiale:
- suprafete articulare acoperite cu cartilaj;
- capsula articulară sau bursă;
- cavitate articulară cu o cantitate mică de lichid sinovial.
Unele articulații au și formațiuni auxiliare sub formă de discuri articulare, menisci și labrum articular.
Suprafețele articulare cel mai adesea corespund între ele în oasele articulate.
Țesutul conjunctiv - structură, funcții, compoziție
Sunt acoperite cu cartilaj articular, care facilitează alunecarea suprafețelor articulare și atenuează șocurile.
Capsulă comună crește până la oasele articulare de-a lungul marginii suprafețelor lor articulare sau retrăgându-se ușor de ele și înconjoară ermetic cavitatea articulară.
Capsula are 2 straturi: fibros exterior si sinovial intern.
Strat fibros pe alocuri formează ligamente - îngroșări care întăresc capsula și acționează și ca frâne pasive, limitând mișcarea în articulație.
Stratul sinovial subţire. Captuseste stratul fibros din interior si continua pe suprafata osului, neacoperit cu cartilaj articular.
Cavitatea articulară Este un spațiu asemănător unei fante, închis ermetic, limitat de suprafețe articulare și de o membrană sinovială. Cavitatea articulară conține o cantitate mică de lichid sinovial.
3. Semiarticulatii sau simfize (hemiartroza) - conexiuni de trecere de la continuu la discontinu sau invers. Aceștia sunt compuși cartilaginoși sau fibroși, în grosimea cărora există o mică cavitate sub formă de gol.
Clasificarea îmbinărilor
În îmbinări, în funcție de structura suprafețelor de articulare (formă, curbură, mărime), se pot produce mișcări în jurul diferitelor axe. În biomecanica articulațiilor se disting următoarele axe de rotație: 1) frontală, 2) sagitală și 3) verticală. În plus, se distinge mișcarea circulară.
Articulațiile sunt clasificate după următoarele criterii:
- după numărul de suprafeţe articulare;
- dupa forma suprafetelor articulare;
- dupa functie.
eu. Pe baza numărului de suprafețe articulare, acestea se disting:
A) articulație simplă– are 2 suprafețe articulare (de ex. humerus, interfalangian)
b) îmbinare compusă– are mai mult de 2 suprafete articulare (ex: cot, genunchi). O articulație complexă este formată din mai multe articulații simple în care mișcările pot fi efectuate separat.
V) articulație complexă– conține cartilaj intra-articular în interiorul capsulei articulare, care împarte articulația în două camere (de exemplu, articulația temporomandibulară, genunchi).
G) îmbinare combinată– reprezintă o combinație de mai multe articulații izolate una de cealaltă, articulații situate separat una de cealaltă, dar funcționând împreună (de exemplu, ambele articulații temporomandibulare, articulații radio-ulnare proximală și distală etc.)
II. După formă și funcție, clasificarea se realizează astfel: funcția unei articulații este determinată de numărul de axe în jurul cărora se fac mișcările. Numărul acestor axe depinde de forma suprafețelor articulare ale articulației. Pe baza acestui fapt, articulațiile se disting:
1. Articulații uniaxiale(cilindric sau rotativ și în formă de bloc):
2. Articulații biaxiale (elipsoid, şa, condilar):
3. Articulații triaxiale sau multiaxiale(sferic, în formă de nucă, plat):
©2015-2018 poisk-ru.ru
Țesuturile conjunctive animale
Cea mai mare parte a masei corporale a animalelor este formată din țesut conjunctiv. Ele constau din cartilaj, oase, tendoane și ligamente.
Caracteristicile țesuturilor conjunctive
Structura țesuturilor conjunctive este diferită la diferite animale și în diferite părți ale aceluiași organism. Mai mult, o caracteristică comună a structurii lor este că celulele par să fie împrăștiate în masa de substanță intercelulară. Există mai multe tipuri de țesuturi conjunctive care îndeplinesc diferite funcții.
Țesut conjunctiv fibros
Țesutul conjunctiv fibros se găsește peste tot în corpul animalului. Conectează pielea cu mușchii, ținând-o în poziția dorită și conectează organele. Celulele acestui tip de țesut sunt înconjurate de o rețea densă de fibre care formează substanța intercelulară.
Os
Țesutul osos formează oasele scheletului - suportul intern al vertebratelor. Țesutul osos este format din substanțe minerale, care îi conferă rezistență, și substanțe organice, care îi conferă elasticitate.
Structura și funcțiile țesutului conjunctiv, principalele tipuri de celule
Această combinație ajută țesutul osos să îndeplinească o funcție de susținere.
Celulele osoase rămân în viață și secretă substanță intercelulară pe toată durata vieții animalului. Celulele sunt conectate între ele prin numeroase procese care se află în substanța interosoasă.
Țesutul osos formează oase. Creșterea și nutriția oaselor formate din țesutul osos este asigurată de periostul care le acoperă.
Țesutul cartilajului
Țesutul cartilajului acoperă capetele oaselor și se găsește la nivelul articulațiilor, ceea ce conferă scheletului flexibilitate.
Celulele țesutului cartilajului, singure sau în grupuri, sunt scufundate într-o substanță intercelulară elastică. Scheletele rechinilor și razelor nu au țesut osos, sunt construite în întregime din cartilaj. La om, cartilajul poate fi simțit în auriculă și în vârful nasului.
Sânge
Sângele este un țesut conjunctiv special. Conține o substanță intercelulară lichidă - plasmă. Plasma conține celule sanguine: eritrocite (globule roșii), leucocite (globule albe) și trombocite (celule rotunde, ovale sau trombocite).
Când sângele se deplasează prin cele mai mici vase - capilare, nutrienții în stare dizolvată pătrund în spațiul intercelular. Ca rezultat, se formează lichid tisular. Din ea ia naștere limfa (gr. limpha - umiditate, apă curată), care se adună în vasele limfatice și din acestea intră din nou în sânge.
Sângele, limfa și lichidul tisular creează mediul intern al corpului.

Țesut adipos
Țesutul adipos este, de asemenea, un țesut conjunctiv. Este format dintr-un număr mare de celule adipoase. Acest țesut este localizat în principal în stratul adipos subcutanat. Stochează grăsimi care pot fi folosite de organism în caz de nutriție insuficientă. În plus, țesutul adipos ajută animalele să rețină căldura și le protejează de șocurile externe.
Caută Prelegeri
Țesut conjunctiv. Localizare în corp, tipuri, structură și funcții.
Constă din celule și o cantitate mare de substanță intercelulară. Substanța intercelulară conține fibre și substanță fundamentală. Fibrele oferă rezistență și elasticitate.
Fibrele sunt împărțite în:
û colagen
û reticular
û elastic
Fibrele de colagen conțin proteina de colagen și sunt foarte durabile.
Fibrele reticulare fac parte din măduva osoasă roșie, ganglionii limfatici și splina. Sunt subțiri și pot forma o rețea fină.
Fibrele elastice conțin proteina elastina, sunt mai puțin durabile decât colagenul și se pot întinde cu ușurință.
Substanța principală care aparține intercelular umple spațiul dintre celule și fibre.
Funcția este variată:
- Țesutul conjunctiv de susținere face parte din oase, cartilaj, ligamente, tendoane și fascia scheletului. Funcția de susținere este îndeplinită de țesut fibros dens (ligamente și tendoane), țesut osos și cartilaj.
- Trofic - această funcție este îndeplinită de sânge și limfă (furnizarea altor țesuturi cu substanțe nutritive).
- Mecanic - țesutul conjunctiv participă la formarea scheletului moale, adică stroma.
- Țesutul conjunctiv este implicat în hematopoieză, adică hematopoieză.
- Țesutul conjunctiv este implicat în fagocitoză.
- Țesutul conjunctiv este implicat în regenerare.
- Funcția respiratorie - participă la procesul de schimb de gaze care are loc în țesuturi și organe.
Țesutul conjunctiv include țesutul conjunctiv în sine, care include fibros lax și fibros dens; țesuturi conjunctive scheletice (cartilaj și os), precum și țesut conjunctiv cu proprietăți speciale (țesut adipos, sânge, limfa și țesuturi hematopoietice).
Țesut conjunctiv fibros lax (FCT).
PBST umple spațiul dintre organe.
PBST include următoarele celule:
Fibroblastele sunt celule plate, în formă de fus.
FUNCȚIILE ȚESUTULUI CONECTIV
Participa la vindecarea rănilor și la formarea țesutului cicatricial.
ü Macrofagele sunt celule care captează și digeră particule străine.
ü Mastocitele produc heparină, care previne coagularea sângelui.
ü Plasma - participă la sinteza anticorpilor.
Anticorpii sunt proteine care protejează împotriva infecțiilor.
ü Celule adipoase – capabile să acumuleze grăsime de rezervă.
ü Celulele pigmentare – conțin granule ale pigmentului melanină.
Țesut conjunctiv fibros dens (DFCT).
În această țesătură, fibrele sunt aranjate strâns. Există puțină substanță intercelulară. PVST face parte din ligamente, tendoane, fascie și membrane.
Fascia este o teacă subțire de țesut conjunctiv în care este plasat un mușchi.
Conține multe fibre de colagen.
Țesutul cartilajului este format din celule condrocite și substanță intercelulară densă.
În substanța intercelulară se găsesc diferite fibre:
ñ hialină
ñ elastic
ñ fibroase
Cartilajul hialin face parte din coaste. Situat la joncțiunea coastei și sternului.
Cartilajul elastic face parte din auriculă și cartilajul laringelui. Calciul nu se depune niciodată în cartilajul elastic.
Cartilajul fibros formează discurile intervertebrale și acoperă articulația maxilarului inferior.
Os.
Constă din celule și substanță intercelulară.
Substanța intercelulară conține substanța principală, care conține multe săruri anorganice (calciu, magneziu).
Substante organice - grasimi, proteine, carbohidrati care contin carbon.
Substante anorganice - saruri minerale.
Datorită acestui fapt, oasele sunt puternice. Osul conține multe săruri de calciu. Dacă nu există suficiente săruri de calciu, se dezvoltă osteoporoza . Osul devine fragil și sunt posibile fracturi.
Dintre sărurile organice din oase, cele mai multe oseina, care conferă oaselor flexibilitate.
Procesul de distrugere și formare de noi celule are loc în mod constant în os.
Există 3 tipuri de celule osoase:
- Osteoblastele sunt celule care formează țesut osos.
- Osteocitele sunt celule care se formează din osteoblaste.
- Osteoclastele sunt celule care distrug țesutul osos.
Există 2 tipuri de țesut osos:
Ø fibră grosieră
Ø placa
Țesutul fibros grosier se găsește la suturile craniului. Constă din fibre de colagen și osteocite.
Țesutul lamelar este mai dens decât țesutul fibros grosier și toate oasele sunt construite din acesta. De asemenea, include un număr mare de fibre de colagen și celule sub formă de plăci.
Unitatea funcțională a osului este osteon.
Țesut adipos
Acesta este țesutul conjunctiv, în care cea mai mare parte este ocupată de celule adipoase - adipocite. Există 2 tipuri: țesut adipos alb (formează acumulări superficiale și profunde), țesut adipos maro (situat între omoplați, în axile, în zona vaselor mari ale gâtului).
Sânge și limfa
Ele constau dintr-o parte lichidă și elemente formate.
©2015-2018 poisk-ru.ru
Toate drepturile aparțin autorilor lor. Acest site nu pretinde autor, dar oferă o utilizare gratuită.
Încălcarea drepturilor de autor și încălcarea datelor cu caracter personal
Țesutul conjunctiv este larg distribuit în organism. Se gaseste in organele interne, in piele, in ligamente, tendoane, membrane ale muschilor si nervilor, in peretele vascular.
Țesutul conjunctiv este format din celule: fibroblaste, histiocite, macrofagocite, bazofile tisulare și substanță intercelulară, care include: fibre - colagen și substanță elastică și fundamentală.
În țesuturi și organe există celule în diferite stadii de formare.
Combinația dintre diferite tipuri de celule și substanță intercelulară determină o mare varietate de structuri și funcții ale țesutului conjunctiv.
Funcțiile țesutului conjunctiv :
1. Trofic (nutriția celulară - sânge, limfa)
2. Protectiv – (fagocitoză, formare de anticorpi)
3. Formează (formează stroma organelor, fascia)
4. Regenerative (regenerarea țesuturilor, vindecarea rănilor)
5. Excretor
Celulele țesutului conjunctiv și substanța intercelulară.
Fibroblastele sunt celule plate, în formă de fus, celulele principale ale țesutului conjunctiv și sunt mobile.
1. Formează substanță intercelulară, sintetizează structuri fibroase:
colagen, elastina, reticulina
2. Capabil de divizare
3. Ele iau naștere din forme slab diferențiate și se pot transforma în alte celule
4. Participa la vindecarea rănilor și la formarea țesutului cicatricial.
Macrofage hepatice - celule stelate
Macrofage pulmonare - fagocite alveolare
Macrofage ale cavităților seroase - macrofage pleurale și peritoneale
țesut osos – osteoblaste
nervos - celule microgliale.
Macrofagocitele - functie principala - fagocitoza - eliminarea din organism a substantelor nocive si straine, celulelor moarte, bacteriilor, virusurilor etc.
Bazofile tisulare(mastocitele) – produc heparină, histamina, serotonină.
Celule plasmatice– produc anticorpi – se găsesc în țesutul conjunctiv lax, mucoasa intestinală, epiploon, ganglioni limfatici și măduva osoasă.
Lipocite– acumulează grăsime de rezervă. Acumularea celulelor adipoase formează țesut adipos.
Celulele reticulare– formează o plasă, în buclele căreia se află celulele țesutului principal.
Celulele adventițiale– situat în stratul exterior al peretelui vaselor de sânge și al organelor goale.
Celulele pigmentare– contin si sintetizeaza boabe de melanina, se gasesc in tesutul conjunctiv, aflat in pielea din jurul anusului, in pielea scrotului si areola glandelor mamare, in coroida ochiului.
Substanța intercelulară a țesutului conjunctiv .
1/ Biopolimerii mucopolizaharidelor B și U - substanță asemănătoare jeleului (heparină, acid hialuronic) - formează rețele și pori
2/ Polimerii glicoproteinelor B și U – se formează în fibroblaste. Formează fibre elastice și de colagen.
3/ Fibre:
Colagen– susține structura specifică a organelor și țesuturilor
Elastic- sub forma unei rețele cu buclă largă.
Reticular– formează o rețea și fac parte din ganglionii limfatici, splina, măduva osoasă etc.
7. Țesut de susținere trofic (conjunctiv adecvat).
ST fibros liber– face parte din vasele de sânge, formează baza formațiunilor limfoide.
Celule: fibroblaste
Există multă substanță intercelulară
Fibre: colagen și elastice – situate difuz, împletite.
ST cu formă fibroasă densă.
Fibrele sunt situate paralel între ele și colectate în mănunchiuri.
Celulele sunt fibroblaste (sunt puține). Țesătura este durabilă, flexibilă și nu se poate întinde. Fibrele sale sunt situate paralel cu liniile de tensiune sub sarcină.
Conținut în sclera, cornee, capsule renale și meninge
ST fibros dens neformat.
Fibrele sunt strâns adiacente între ele - împletite
Există puține celule și substanță fundamentală. Acest tesut este format din: - ligamente
Pereții arterelor
Țesut conjunctiv cu proprietăți speciale– țesut reticular
Compozitie: celulele - reticulocite - sunt capabile sa se transforme in fibroblaste si macrofage.
Fibrele sunt reticulare, formând plexuri complexe.
RT este baza măduvei osoase și a țesutului mieloid.
Țesutul reticular face parte din amigdale și formează membranele mucoase ale intestinelor.
Țesutul cartilajului – se referă la susținerea țesutului conjunctiv împreună cu țesutul osos.
Structura țesutului cartilajului:
1. Celulele– condroblaste și condrocite – situate individual și în grupuri
2. Substanță intercelulară:
a) substanţa principală este densă
b) fibre – colagen (sunt mai multe)
elastic
Tipuri de țesut cartilaj(în funcție de structura substanței intercelulare)
1/ Țesut cartilaj hialin(cartilajul vitros) – conține mai multă substanță de bază. HCT este format din: - schelet embrionar
Suprafețele articulare ale oaselor
Partea cartilaginoasă a coastelor
2/ HT elastic– formată din hialină. Contine mai multe fibre elastice. Cartilajul elastic este baza auriculei, a cartilajului laringelui și a pereților canalului auditiv extern. Este mai puțin transparent, de culoare gălbuie și este capabil să-și refacă forma.
3/ CT fibros– fibrele de colagen sunt colectate în mănunchiuri și ordonate.
VXT este mai puternic, dar mai puțin flexibil decât GC. Formează simfiza pubiană și discurile intervertebrale.
Țesutul fibros al cartilajului funcționează ca un amortizor de șoc.
Cartilajul îndeplinește o funcție mecanică de susținere.
Os - tesutul conjunctiv de sustinere, impreuna cu tesutul cartilaginos, formeaza oasele scheletului.
Structura țesutului osos:
Osteoblastele – formează substanța principală a țesutului osos, sunt situate în zonele de formare osoasă (pe suprafața osului în creștere)
Osteoclaste - celule mari multinucleate cu procese. Participă la distrugerea oaselor și cartilajului, cu formarea de golfuri sau lacune (osul este resorbit datorită enzimelor).
Osteocite – sunt formate din osteoblaste. Au lăstari. Corpurile lor se află în cavitățile osoase, iar procesele lor intră în canaliculele osoase.
Substanța intercelulară este mineralizată.
Fibrele de colagen (fibre de osseină) – conferă oaselor flexibilitate și plasticitate.
Tipuri de țesut osos:
1. Fibroasă grosieră- la embrion si la om la suturile craniului si la locurile de atasare a tendoanelor de oase.
Fibrele de colagen de osseină din țesutul osos grosier formează mănunchiuri, între care osteocitele se află în cavitățile osoase.
2. Lamelar(fibre fine) - toate oasele scheletului.
Fibrele de colagen sunt situate în mănunchiuri paralele în interiorul plăcilor sau între ele.
3. Dentină– celulele sale odontoblastice se află în afara dentinei (corpului), iar procesele lor trec în tubii din interiorul dentinei.
Se formează țesut lamelar
1. Substanță osoasă compactă
2. Substanță osoasă spongioasă alcătuiesc os
În substanță osoasă compactă, plăcile sunt dispuse într-o ordine specială și dau densitatea osoasă (diafize osoase)
În substanța osoasă spongioasă, plăcile formează bare transversale (epifize, oase scurte).
În substanța osoasă compactă, plăcile osoase formează sisteme tubulare deosebite - osteoni (unități structurale ale osului).
Plăcile osoase sunt dispuse concentric în jurul canalului Havers (cavitatea din centrul osteonului), pe unde trece vasul.
Osteocitele sunt situate între plăcile osoase.
periost (periost)– o membrană de țesut conjunctiv format din două straturi.
Stratul exterior este format din țesut conjunctiv mai dens; tendoanele și ligamentele musculare sunt atașate de el.
Stratul interior este format din colagen și fibre elastice, osteoblaste și
osteoclaste.
În timpul creșterii osoase, osteoblastele participă la formarea osului. Periostul conține un număr mare de vase și nervi care pătrund în os și îl hrănesc. Regenerarea osoasă în timpul fracturilor are loc datorită periostului, care, crescând peste locul fracturii, conectează capetele osului rupt, formând un cuplare de țesut osos în jurul lor - calus.
Endost– membrana care acoperă osul din partea canalului medular.
Muşchi.
Anticii spuneau: „Viața este în mișcare”.
Ai întors capul, ai clipit, ai inspirat, te-ai uitat în depărtare, ai spus ceva. În fiecare minut, mii de fibre musculare și celule se contractă în corpul tău. Adăugați la aceasta că inima bate, stomacul bubuie, ureterul transportă ușor urina de la rinichi la vezică, iar vasele de sânge mențin constant o anumită tensiune arterială.
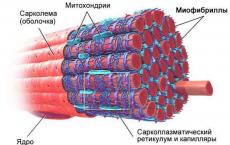
Procesele motorii din corpul uman sunt cauzate de contracția țesutului muscular, care are o proprietate specifică - contractilitatea.
Histologii evidențiază 3 tipuri de tesut muscular:
1. Țesutul muscular neted.
2. Țesut muscular scheletic cu dungi încrucișate.
3. Țesut muscular cardiac striat.
Capitolul 8. ȚESUTUL CONECTIV
Capitolul 8. ȚESUTUL CONECTIV
Tesuturile conjunctive sunt grup marețesuturile derivate din mezenchim, de regulă, sunt polidiferite și cu predominanța substanței intercelulare (structuri fibroase și o componentă amorfă), participând la menținerea constantă a compoziției mediului intern al corpului.
Țesutul conjunctiv reprezintă mai mult de 50% din greutatea corpului uman. Participă la formarea stromei organelor, a straturilor dintre alte țesuturi, a dermei pielii și a scheletului. Natura multifuncțională a țesuturilor conjunctive este determinată de complexitatea compoziției și organizării lor.
Funcțiile țesuturilor conjunctive. Tesuturile conjunctive indeplinesc diverse functii: trofice, protectoare, de sustinere (biomecanice), plastice, morfogenetice. Funcția trofică în sensul larg al cuvântului este asociată cu reglarea nutriției diferitelor structuri tisulare, cu participarea la metabolism și menținerea constantă a compoziției mediului intern al corpului. În implementarea acestei funcții, rolul principal îl joacă substanța principală prin care se realizează transportul apei, sărurilor și moleculelor nutritive - mediul tampon integrator. Funcția de protecție este de a proteja organismul de influențe mecanice nefiziologice (deteriorări) și de a neutraliza substanțele străine venite din exterior sau formate în interiorul corpului. Aceasta este asigurată de protecția fizică (țesutul osos), precum și de activitatea fagocitară a macrofagelor și a celulelor imunocompetente implicate în reacțiile imunității celulare și umorale. Funcția de susținere (biomecanică) este asigurată în primul rând de fibrele de colagen și elastice, care formează fundațiile fibroase ale tuturor organelor, și de compoziția și proprietățile fizico-chimice ale substanței intercelulare a țesuturilor scheletice (mineralizare). Cu cât substanța intercelulară este mai densă, cu atât funcția biomecanică de susținere este mai semnificativă. Funcția plastică a țesutului conjunctiv se exprimă prin adaptarea la condițiile de viață în schimbare, regenerare și participarea la înlocuirea defectelor organelor atunci când acestea sunt deteriorate. Morfogenetic
Funcția (formatoare de structură) se manifestă prin formarea complexelor tisulare și asigurarea organizării structurale generale a organelor (formarea de capsule, septuri intraorgane), influența reglatoare a unora dintre componentele sale asupra proliferării și diferențierii celulelor diferitelor țesuturi.
Clasificarea țesuturilor conjunctive. Tipurile de țesut conjunctiv diferă în compoziția și raportul diferențelor celulare, fibrelor, precum și proprietati fizice si chimice substanță intercelulară amorfă. Țesuturile conjunctive sunt împărțite în țesut conjunctiv propriu-zis (țesut conjunctiv lax și țesuturi conjunctive cu proprietăți speciale) și țesuturi scheletice. Acestea din urmă, la rândul lor, sunt împărțite în trei tipuri de țesut cartilaginos (hialin, elastic, fibros), două tipuri de țesut osos (reticulofibros și lamelar), precum și ciment și dentina dintelui (Diagrama 8.1).
Histogenia țesuturilor conjunctive. Sursa dezvoltării țesuturilor conjunctive este mezenchimul (din greacă. mesos- in medie, enchima- masa de umplere). Acesta este unul dintre rudimentele embrionare (după unele idei - țesut embrionar), care este o parte slăbită a stratului germinal mijlociu. Elementele celulare ale mezenchimului se formează în procesul de diferențiere a dermatomului, sclerotomului, straturilor viscerale și parietale ale splanhnotomului. În plus, există ectomezenchim (neuromezenchim), care se dezvoltă din creasta neură (placa ganglionară). Pe măsură ce embrionul se dezvoltă, celulele de origine diferită de alte celule embrionare migrează în mezenchim.
Schema 8.1. Clasificarea țesuturilor conjunctive
rudimente, de exemplu, celulele neuroblastice diferite, mioblastele din angajarea mușchilor scheletici, pigmentocitele etc.
În consecință, dintr-un anumit stadiu de dezvoltare a embrionului, mezenchimul este un mozaic de celule care provin din diferite straturi germinale și rudimente de țesut embrionar. Cu toate acestea, din punct de vedere morfologic, toate celulele mezenchimale diferă puțin unele de altele și numai metodele de cercetare foarte sensibile (de regulă, imunocitochimice, microscopice electronice) dezvăluie celule de natură diferită în interiorul mezenchimului. Mezenchimul există doar în perioada embrionară a dezvoltării umane. După naștere, în corpul uman rămân doar celule slab diferențiate (pluripotente) ca parte a țesutului conjunctiv fibros lax (celule adventițiale), care se pot diferenția divergent în direcții diferite, dar într-un anumit sistem de țesuturi.
Există histogeneză embrionară și postembrionară a țesuturilor conjunctive. În procesul de histogeneză embrionară, mezenchimul dobândește caracteristicile structurii tisulare înainte de formarea altor țesuturi. Acest proces are loc diferit în diferite organe și sisteme și depinde de semnificația lor fiziologică inegală în diferite stadii ale embriogenezei. În diferențierea mezenchimului se remarcă asincronia topografică atât la nivelul embrionului, cât și la nivelul organelor extraembrionare, rate mari de reproducere celulară și formare de fibre. Histogenia postembrionară în condiții fiziologice normale are loc mai lent și are ca scop menținerea homeostaziei tisulare, proliferarea celulelor slab diferențiate și înlocuirea celulelor muribunde. Un rol semnificativ în aceste procese îl joacă interacțiunile intrațesute intercelulare, factori inductori și inhibitori (integrine, factori adezivi intercelulari, încărcări funcționale, hormoni, oxigenare, prezența celulelor slab diferențiate).
Principii generale de organizare a țesutului conjunctiv. Componentele principale ale țesuturilor conjunctive sunt derivații celulari - structuri fibroase de colagen și tipuri elastice, o substanță de bază (amorfă) care joacă rolul unui mediu metabolic tampon integrator și elementele celulare care creează și mențin raportul cantitativ și calitativ al compoziției. a componentelor necelulare.
Specificul de organ al elementelor celulare ale țesutului conjunctiv este exprimat în cantitate, formă și raport diferite feluri celulele, metabolismul și funcțiile acestora, adaptate optim la funcția organului. Specificul elementelor celulare se manifestă și prin interacțiunea lor între ele (localizate individual, asociații celulare), caracteristicile lor. structura interna(compoziția organelor, structura nucleară, prezența enzimelor etc.). Specificitatea țesutului conjunctiv este, de asemenea, relevată în raportul dintre celulele diferitelor difere și structurile necelulare din diferite părți ale corpului. În țesutul conjunctiv fibros lax, celulele diferitelor difere și materie amorfă prevalează asupra fibrelor, iar în țesutul conjunctiv dens, dimpotrivă, cea mai mare parte a masei sale este formată din fibre.
8.1. ȚESUT CONECTIV CORRECT
8.1.1. Țesuturile conjunctive fibroase
Țesut conjunctiv lax
Țesut conjunctiv lax (textus connectivus laxus) se găsește în toate organele, deoarece însoțește vasele de sânge și limfatice și formează stroma multor organe. În ciuda prezenței caracteristicilor organelor, structura țesutului conjunctiv lax din diferite organe este similară. Este alcătuită din celule de diferite determinari histogenetice și substanțe intercelulare (Fig. 8.1).

Orez. 8.1.Țesut conjunctiv lax:
A- prepararea peliculei: 1 - fibroblast; 2 - macrofage; 3 - fibre de colagen; 4 - fibre elastice; 5 - limfocit; b- fibroblast; V- macrofage

Orez. 8.2. Structura ultramicroscopică a unui fibroblast în diferite stadii de diferențiere (după N. A. Yurina și A. I. Radostina, cu modificări): Fibroblaste: A- slab diferenţiat; b- tineri; V- matur; G- fibrocite. 1 - miez; 2 - complexul Golgi; 3 - mitocondrii; 4 - ribozomi și poliribozomi; 5 - reticul endoplasmatic granular; 6 - fibrile de colagen
Celulele
Principalele celule ale țesutului conjunctiv sunt fibroblaste(familie de celule formatoare de fibrile), macrofage(familie), mastocite, celule adventice, plasmocite, pericite, celule adipoase,și leucocite, migrează din sânge; Uneori pigment
celule.
Fibroblastele(din lat. fibră- fibre, greacă blastos- germinare, rudiment) - celule care sintetizeaza componentele substantei intercelulare: proteine (colagen, elastina), proteoglicani, glicoproteine (vezi Fig. 8.1; Fig. 8.2, 8.3).
Printre celulele mezenchimale se numără celule stem care dau naștere diferențierii fibroblastelor: celule stem, celule semi-stem precursoare, slab diferențiate (prost specializate), fibroblaste diferențiate (mature, care funcționează activ), fibrocite (definitive - formele finale de celule), precum și miofibroblaste și fibroclaste. Funcția principală a fibroblastelor este asociată cu formarea substanței și fibrelor principale, vindecarea rănilor, dezvoltarea țesutului de granulație, formarea unei capsule de țesut conjunctiv în jurul unui corp străin etc. Din punct de vedere morfologic, în acest diferențial pot fi identificate doar celule, începând cu fibroblast slab diferențiat. Acestea din urmă sunt celule cu proces mic, cu nucleul rotund sau oval și cu un nucleol mic, citoplasmă bazofilă, bogată în ARN. Dimensiunea celulei nu depășește 20-25 microni. În citoplasma acestor celule se găsește
un număr mare de ribozomi liberi (vezi Fig. 8.2). Reticulul endoplasmatic și mitocondriile sunt slab dezvoltate. Complexul Golgi este reprezentat de grupuri de tuburi scurte și vezicule.
Fibroblaste mature diferențiate dimensiuni mai mari și răspândite pe preparate de film poate ajunge la 40-50 microni sau mai mult (vezi Fig. 8.1). Acestea sunt celule care funcționează activ. Nucleii lor sunt usori, ovali, contin 1-2 nucleoli mari; citoplasma este bazofilă, cu un reticul endoplasmatic granular bine dezvoltat, care pe alocuri este în contact cu plasmalema (vezi Fig. 8.2, Fig. 8.3, A). Complexul Golgi este distribuit sub formă de cisterne și vezicule în întreaga celulă. Mitocondriile și lizozomii sunt moderat dezvoltați.
În citoplasma fibroblastelor, în special în partea periferică, există microfilamente cu grosimea de 5-6 nm care conțin proteine precum actina și miozina, ceea ce determină capacitatea acestor celule de a se deplasa. Mișcarea fibroblastelor devine posibilă numai după ce se leagă de structurile fibrilare de susținere (fibrină, fibre de țesut conjunctiv) cu ajutorul fibronectinei, glicoproteină sintetizată de fibroblaste și alte celule care asigură aderența celulelor și a structurilor necelulare. În timpul mișcării, fibroblastul devine aplatizat, iar suprafața sa poate crește de 10 ori.
Plasmalema fibroblastelor este o zonă importantă de receptor care mediază efectele diferiților factori de reglare. Activarea fibroblastelor este de obicei însoțită de acumularea de glicogen și creșterea activității enzimelor hidrolitice. Energia generată de metabolismul glicogenului este utilizată pentru a sintetiza polipeptide și alte componente secretate de celulă.
Biosinteza proteinelor de colagen și elastină, proteoglicani, necesare formării substanței principale și a fibrelor, în fibroblastele mature se realizează destul de intens, mai ales în condiții de concentrație scăzută de oxigen. Factorii de stimulare a biosintezei colagenului sunt fierul, cuprul, ionii de crom și acidul ascorbic. Una dintre enzimele hidrolitice, colagenaza, descompune colagenul imatur din interiorul celulelor, care aparent reglează intensitatea secreției de colagen la nivel celular.
Pe baza capacității lor de a sintetiza proteine fibrilare, familia fibroblastelor include celule reticulare ale țesutului conjunctiv reticular al organelor hematopoietice, condroblaste și osteoblaste ale tipurilor scheletice de țesut conjunctiv.
Fibrocite- forme definitive de dezvoltare a fibroblastelor. Aceste celule sunt în formă de fus cu procese în formă de aripi. Conțin un număr mic de organite, vacuole, lipide și glicogen. Sinteza colagenului și a altor substanțe din fibrocite este redusă brusc.
Miofibroblaste- celule asemănătoare morfologic cu fibroblastele, combinând capacitatea de a sintetiza nu numai colagen, ci și contractilitatea

Orez. 8.3. Fibroblast, miofibroblast și fibroclaste:
A- fibroblast (preparat de A. I. Radostina): 1 - nucleu; 2 - reticul endoplasmatic granular; 3 - ribozomi; 4 - complexul Golgi; 5 - mitocondrii; 6 - plasma-molema; 7 - fibrile de colagen; b- miofibroblast (preparat A. B. Schechter): 1 - nucleu; 2 - reticul endoplasmatic granular; 3 - ribozomi; 4 - complexul Golgi; 5 - filamente contractile; 6 - plasmalema
proteinele corpului în cantități semnificative (vezi Fig. 8.3, b). S-a stabilit că fibroblastele se pot dezvolta în miofibroblaste, care sunt similare din punct de vedere funcțional cu celulele musculare netede, dar spre deosebire de acestea din urmă au un reticul endoplasmatic bine dezvoltat. Astfel de celule sunt observate în

Orez. 8.3. Continuare:
V- fibroclaste (preparat de A. B. Shekhter): 1 - miez; 2 - reticul endoplasmatic granular; 3 - ribozomi; 4 - lizozomi; 5 - fagolizozomi cu fragmente de fibrile de colagen. Microfotografii electronice. Mărire 20.000
țesut de granulație în condiții de regenerare a procesului plăgii și în uter în timpul dezvoltării sarcinii.
Fibroclaste- celulele cu activitate fagocitară și hidrolitică ridicată, participă la „resorbția” substanței intercelulare (vezi Fig. 8.3, c) în timpul perioadei de involuție a organelor (de exemplu, uterul după sfârșitul sarcinii). Ele combină caracteristicile structurale ale celulelor formatoare de fibrile (reticul endoplasmatic granular dezvoltat, complex Golgi, mitocondrii relativ mari, dar puține), precum și lizozomi cu enzimele lor hidrolitice caracteristice. Complexul de enzime pe care le secretă în afara celulei descompune substanța de cimentare a fibrelor de colagen, după care are loc fagocitoza și digestia intracelulară a colagenului de către proteazele acide ale lizozomilor.
Macrofage(macrofagocite) (din greacă. makros- mare, lung, fagos- devoratoare) este o populatie eterogena de celule specializate a sistemului de aparare al organismului. Există două grupuri de macrofage - libere și fixe. Macrofagele libere includ macrofagele de țesut conjunctiv lax, sau histiocite; macrofagele cavităților seroase; macrofagele exsudatelor inflamatorii; macrofagele alveolare ale plămânilor. Macrofagele sunt capabile să se miște în tot corpul. Grupul de macrofage fixe (rezidente) este format din macrofage ale măduvei osoase, țesutului osos și cartilajului (osteoclaste, condroclaste),

Orez. 8.4. Macrofage:
A- macrofage ale țesutului conjunctiv subcutanat al șobolanului (micrograf, colorat cu hematoxilină de fier); b- macrofage (micrografia electronică; preparat de A.I. Radostina, mărire 18.000): 1 - nucleu; 2 - lizozomi primari; 3 - lizozomi secundari; 4 - profilele tubilor reticulului endoplasmatic; 5 - microcreșteri ale stratului periferic al citoplasmei
zenks, ganglioni limfatici (macrofage dendritice), macrofage intraepidermice (celule Langerhans), macrofage vilozități placentare (celule Hofbauer), macrofage ale sistemului nervos central (microglia).
Mărimea și forma macrofagelor variază în funcție de starea lor funcțională (Fig. 8.4). De obicei macrofage, cu excepția unora dintre ele
speciile (celule gigantice ale corpurilor străine, condro- și osteoclaste), au un singur nucleu. Nucleii macrofagelor sunt mici, rotunzi, în formă de fasole sau neregulate. Conțin aglomerări mari de cromatină. Citoplasma este bazofilă, bogată în lizozomi, fagozomi (trăsături distinctive) și vezicule pinocitotice, conține o cantitate moderată de mitocondrii, reticul endoplasmatic granular, complex Golgi, incluziuni de glicogen, lipide și altele (vezi Fig. 8.4, b). În citoplasma macrofagelor, se distinge o „periferie celulară”, care oferă macrofagului capacitatea de a se mișca, de a atrage microprotruzii ale citoplasmei și de a efectua endo- și exocitoză. Direct sub plasmalemă există o rețea de filamente de actină cu un diametru de 5-6 nm. Microtubulii cu un diametru de 20 nm trec prin această rețea și se atașează de plasmalemă. Microtubulii sunt direcționați radial de la centrul celulei către periferia celulei și joacă un rol important în mișcările intracelulare ale lizozomilor, veziculelor micropinocitotice și altor structuri. Pe suprafața membranei plasmatice există receptori pentru celulele tumorale și celulele roșii din sânge, limfocitele T și B, antigenele, imunoglobulinele și hormonii. Prezența receptorilor de imunoglobuline determină participarea macrofagelor la reacțiile imune (vezi capitolul 14).
Forme de manifestare a funcției de protecție a macrofagelor: 1) absorbția și descompunerea ulterioară sau izolarea materialului străin; 2) neutralizarea acestuia la contact direct; 3) transferul de informații despre material străin către celule imunocompetente capabile să-l neutralizeze; 4) oferind un efect stimulativ asupra unei alte populații de celule a sistemului de apărare al organismului. Macrofagele au organele care sintetizează enzime pentru descompunerea intracelulară și extracelulară a materialelor străine, antibacteriene și alte substanțe biologic active (proteaze, hidrolaze acide, pirogen, interferon, lizozim etc.).
Numărul de macrofage și activitatea lor cresc în special odată cu procese inflamatorii. Macrofagele produc factori chemotactici pentru leucocite. IL-1 secretată de macrofage este capabilă să mărească aderența leucocitelor la endoteliu, secreția de enzime lizozomale de către neutrofile și citotoxicitatea acestora și activează sinteza ADN-ului în limfocite. Macrofagele produc factori care activează producerea de imunoglobuline de către limfocitele B și diferențierea limfocitelor T și B; factorii antitumorali citolitici, precum și factorii de creștere care influențează reproducerea și diferențierea celulelor propriei populații, stimulează funcția fibroblastelor (vezi capitolul 14).
Contactul macrofagelor cu antigenele crește brusc consumul de glucoză, metabolismul lipidic și activitatea fagocitară.
Macrofagele sunt formate din HSC, precum și din promonocite și monocite (vezi Fig. 7.15). Reînnoirea completă a macrofagelor țesutului conjunctiv fibros laxe la animalele de experiment are loc de aproximativ 10 ori mai rapid decât fibroblastele.
Unul dintre tipurile de macrofage sunt celulele gigantice multinucleate, care anterior erau numite „celule gigantice străine”.
„corpuri”, deoarece se pot forma, în special, în prezența unui corp străin. Celulele gigantice multinucleate conțin 10-20 de nuclei sau mai mulți și apar fie prin fuziunea macrofagelor mononucleare, fie prin endomitoză fără citotomie. Conform microscopiei electronice, celulele gigantice multinucleate conțin un aparat sintetic și secretor și un număr mare de lizozomi. Plasmalema formează numeroase pliuri.
Conceptul de sistem macrofage. Acest sistem include macrofage (histiocite) ale țesutului conjunctiv fibros lax, celule stelate ale vaselor sinusoidale ale ficatului, macrofage libere și fixe ale organelor hematopoietice (măduvă osoasă, splină, ganglioni limfatici), macrofage ale plămânilor, exsudate inflamatorii (peritoneale). macrofage), osteoclaste, celule gigantice ale corpurilor străine.corpi și macrofage gliale ale țesutului nervos (microglia). Toate sunt fagocite active. Materialul fagocitat suferă clivaj enzimatic în interiorul celulei („fagocitoză completă”), datorită căruia sunt eliminați agenții nocivi pentru organism care apar local sau pătrund din exterior. Celulele au receptori de imunoglobuline pe suprafața lor și provin din promonocitele măduvei osoase și monocitele din sânge. Spre deosebire de astfel de fagocitele „profesionale”, fibroblastele, celulele reticulare, celulele endoteliale, granulocitele neutrofile etc. au capacitatea de absorbție facultativă, dar aceste celule nu fac parte din sistemul macrofagic.
I. I. Mechnikov a fost primul care a venit la ideea că fagocitoza, care apare în procesul de evoluție ca formă de digestie intracelulară și este atribuită multor celule, este în același timp un mecanism de protecție important. El a fundamentat fezabilitatea combinării lor într-un singur sistem și a propus numirea acestuia macrofage. Sistemul de macrofage este un aparat de protecție puternic care participă atât la reacțiile de protecție generale, cât și la cele locale ale corpului. În întregul organism, sistemul macrofagelor este reglat atât prin mecanisme locale, cât și prin sistemele nervos și endocrin.
În anii 1930-1940. acest sistem protector a fost numit reticuloendotelial. ÎN În ultima vreme se numeste sistemul fagocitelor mononucleare, care insa nu il caracterizeaza cu acuratete datorita faptului ca printre celulele incluse in acest sistem se numara si cele multinucleate (osteoclaste).
Mastocitele(mastocite, bazofile tisulare, mastocite). Acești termeni se referă la celulele din citoplasmă ale cărora există o granularitate specifică, care amintește de granulele granulocitelor bazofile. Mastocitele participă la reducerea coagulării sângelui, la creșterea permeabilității barierei sânge-țesut, în procesul de inflamație, imuno-
geneza etc. La om, mastocitele se găsesc oriunde există straturi de țesut conjunctiv lax. Există în special multe mastocite în peretele tractului digestiv, uter, glanda mamară, timus și amigdale. Ele sunt adesea localizate în grupuri de-a lungul vaselor de sânge ale microvasculaturii - capilare, arteriole, venule și vase limfatice mici (Fig. 8.5, A).

Orez. 8.5. Mastocitele: A -în țesutul conjunctiv subcutanat (micrograf): 1 - nucleu; 2 - granule metacromatice în citoplasmă; b- diagrama structurii ultramicroscopice (conform lui Yu. I. Afanasyev): 1 - miez; 2 - complexul Golgi; 3 - lizozom; 4 - mitocondrii; 5 - reticul endoplasmatic; 6 - microvilozități; 7 - granule eterogene; 8 - granule secretoare în substanţa intercelulară
Forma mastocitelor este variată. Celulele pot avea formă neregulată, ovale. Uneori, aceste celule au procese scurte și largi, ceea ce se datorează capacității lor de a mișca ameboide. La om, lățimea acestor celule variază de la 4 la 14 microni, lungimea de până la 22 microni. Nucleii celulari sunt relativ mici, de obicei de formă rotundă sau ovală, cu cromatina dens. Există numeroase granule în citoplasmă. Mărimea, compoziția și cantitatea granulelor variază. Diametrul lor este de aproximativ 0,3-1 microni (Fig. 8.5, b). O minoritate de granule sunt lizozomi azurofili care colorează ortocromatic. Majoritatea granulelor sunt caracterizate prin metacromazie și conțin heparină, acizi condroitinsulfuric de tip A și C, acid hialuronic, histamina și serotonină. Granulele au structura reticulata, lamelara, cristaloida si mixta.
Organelele mastocitelor (mitocondriile, complexul Golgi, reticulul endoplasmatic) sunt slab dezvoltate. În citoplasmă se găsesc diferite enzime: proteaze, lipaze, fosfataze acide și alcaline, peroxidază, citocrom oxidază, ATPaza etc. Totuși, histidin decarboxilaza, care este folosită pentru a sintetiza histaminei din histidină, ar trebui considerată o enzimă marker.
Mastocitele sunt capabile să secrete și să elibereze granulele lor. Degranularea mastocitelor poate apărea ca răspuns la orice modificare a condițiilor fiziologice și a acțiunii agenților patogeni. Cu toate acestea, eliberarea de amine biogene din mastocit poate avea loc și prin secreția de componente solubile (secreția de histamină). Histamina determină imediat dilatarea capilarelor sanguine și crește permeabilitatea acestora, care se manifestă prin edem local. De asemenea, are proprietăți hipotensive și este un mediator important al inflamației.
Histamina acționează ca un antagonist al heparinei, care reduce permeabilitatea substanței intercelulare și coagularea sângelui și are un efect antiinflamator.
Numărul de mastocite se modifică în funcție de condițiile fiziologice ale corpului: crește în uter și glandele mamare în timpul sarcinii și în stomac, intestine și ficat la apogeul digestiei.
Precursorii mastocitelor provin din celulele stem hematopoietice ale măduvei osoase roșii. Procesele de diviziune mitotică a mastocitelor sunt observate extrem de rar.
Celule plasmatice (plasmocite). Aceste celule asigură producerea de anticorpi – gama globuline (proteine) atunci când în organism apare un antigen. Ele se formează în organele limfoide din limfocitele B (vezi capitolul 14), întâlnite de obicei în țesutul conjunctiv lax al laminei propria a membranelor mucoase ale organelor goale, epiploon, țesutul conjunctiv interstițial al diferitelor glande (mamare, salivare etc. ), ganglioni limfatici, splină, măduvă osoasă etc.
Dimensiunea celulelor plasmatice variază de la 7 la 10 microni. Forma celulelor este rotundă sau ovală. Sâmburii sunt relativ mici, de formă rotundă sau ovală și sunt situate excentric. Citoplasma este puternic bazofilă, conține endoplasmă granulară bine dezvoltată, localizată concentric

Orez. 8.6. Adipocitele:
1 - capilar; 2 - incluziuni lipidice; 3 - miez; 4 - citoplasmă (micrograf). Colorare - hematoxilină de fier
rețea tică în care se sintetizează proteinele (anticorpii). Bazofilia este absentă numai într-o zonă mică de lumină a citoplasmei în apropierea nucleului, formând așa-numita sferă sau curte. Aici se află centriolii și complexul Golgi. Uneori, acumulări de imunoglobuline se găsesc sub formă de oxifil corpusculul lui Roussel.
Celulele plasmatice se caracterizează printr-o rată ridicată de sinteză și secreție de anticorpi, ceea ce le diferențiază de predecesorii lor. Un aparat secretor bine dezvoltat permite sinteza și secreția a câteva mii de molecule de imunoglobuline pe secundă. Numărul de celule plasmatice crește în diferite boli infecțioase, alergice și inflamatorii.
Adipocite (celule adipoase). Acesta este numele pentru celulele care au capacitatea de a acumula cantități mari de grăsime de rezervă, care participă la trofism, producerea de energie și metabolismul apei. Adipocitele sunt localizate în grupuri, mai rar individual și, de regulă, în apropierea vaselor de sânge. Acumulând în cantități mari, aceste celule formează țesut adipos (alb sau maro) (vezi mai jos).
Forma celulelor adipoase simple este sferică. O celulă de grăsime matură conține de obicei o picătură mare de grăsime neutră (trigliceride), ocupând toată partea centrală a celulei și înconjurată de o margine citoplasmatică subțire, în a cărei parte îngroșată se află nucleul (Fig. 8.6). În plus, citoplasma adipocitelor conține o cantitate mică de alte lipide: colesterol, fosfolipide, acizi grași liberi etc. Lipidele sunt bine colorate cu Sudan III în portocaliu sau cu acid osmic în negru. În citoplasma adiacentă nucleului și uneori în partea opusă mai subțire a acestuia

Orez. 8.7. Pigmentocite în piele (micrograf). Preparatul nu este colorat
sunt dezvăluite mitocondrii în formă de baston și filamentoase cu cresta dens situată. La periferia celulei se găsesc numeroase vezicule pinocitotice.
Adipocitele din țesutul adipos maro se disting prin incluziuni lipidice mici și mitocondriile situate în jurul nucleului. Adipocitele au o mare capacitate metabolică. Atât numărul de incluziuni grase din adipocite, cât și numărul de celule adipoase în sine din țesutul conjunctiv lax sunt supuse unor fluctuații semnificative.
Funcțiile adipocitelor sunt trofice, asociate cu furnizarea de energie și rezerve de apă în organism, precum și cu participarea la procesul de termoreglare.
Noi celule adipoase din țesutul conjunctiv al corpului adult se pot dezvolta cu o nutriție sporită din celulele adventițiale (cambiale) adiacente capilarelor sanguine. În acest caz, în citoplasma celulelor apar mai întâi picături mici de grăsime, care, crescând în dimensiune, se contopesc treptat în picături mai mari. Pe măsură ce picătura de grăsime crește, reticulul endoplasmatic și complexul Golgi sunt reduse, iar nucleul este comprimat, aplatizat și împins la periferia celulei.
Grupurile de celule adipoase găsite în multe organe sunt numite țesut adipos.
Celulele adventițiale. Acestea sunt celule slab diferențiate (cambiale) care însoțesc vasele de sânge mici. Au o formă turtită sau fusiformă, cu citoplasmă ușor bazofilă, un nucleu oval și un număr mic de organele. Aceste celule, prin diferențiere divergentă, dau naștere la diferite diferențieri celulare (fibroblastice, miofibroblastice, adipocite etc.).
Pericitele- celulele care înconjoară capilarele sanguine și alcătuiesc pereții acestora (vezi capitolul 13).
Pigmentocite(celule pigmentare, melanocite). Aceste celule conțin pigmentul melanină în citoplasmă. Sunt mulți dintre ei în semne de nastere, precum și în țesutul conjunctiv al oamenilor din rasele negre și galbene. Pigmentocitele au procese scurte, de formă neregulată (Fig. 8.7), un număr mare de melanozomi (granule de melanină) măsurând 15-25 nm și ribozomi. Unii melanozomi din melanocite migrează către keratinocitele straturilor spinoase și bazale ale epidermei.
Citoplasma melanocitelor conține și amine biologic active, care pot participa, împreună cu mastocitele, la reglarea tonusului pereților vasculari (vezi capitolul 14).
Melanocitele aparțin doar formal țesutului conjunctiv, deoarece sunt localizate în acesta. În ceea ce privește originea lor, s-a dovedit formarea acestor celule din celule stem din crestele neurale, și nu din mezenchim. Celulele țesutului conjunctiv sunt conectate funcțional într-un singur sistem datorită numeroșilor factori de interacțiune, în special în procesele de inflamație și regenerare post-traumatică, cu încălcarea regimului de apă-sare al organismului etc.
Substanță intercelulară
Substanță intercelulară, sau matrice (substanța intercellularis),Țesutul conjunctiv este format din colagen și fibre elastice, precum și substanță măcinată (amorfă). Substanța intercelulară atât la embrioni, cât și la adulți este formată, pe de o parte, prin secreția efectuată de celulele țesutului conjunctiv și, pe de altă parte, din plasma sanguină care pătrunde în spațiile intercelulare.
La embrionii umani, formarea substanței intercelulare are loc începând cu luna 1-2 de dezvoltare intrauterină. În timpul vieții, substanța intercelulară este în mod constant reînnoită.
Structurile de colagen care alcătuiesc țesuturile conjunctive ale organismelor umane și animale sunt componentele sale cele mai reprezentative, formând o ierarhie organizațională complexă. Baza întregului grup de structuri de colagen este proteina fibroasă - colagen, care determină proprietăţile structurilor de colagen.
Fibre de colagen (fibrae collagenae)în compoziția diferitelor tipuri de țesut conjunctiv determină rezistența acestora. În țesutul conjunctiv lax, neformat, acestea sunt localizate în direcții diferite sub formă de fire ondulate curbate, răsucite în spirală, rotunde sau aplatizate, cu o grosime de 1-3 microni sau mai mult. Lungimea lor este diferită. Structura internă a fibrei de colagen este determinată de proteina fibrilă - colagen, care este sintetizat pe ribozomii reticulului endoplasmatic granular al fibroblastelor.
Există aproximativ 20 de tipuri de colagen, care diferă în organizarea moleculară, afilierea organelor și țesuturilor.

Orez. 8.8. Biosinteza și fibrilogeneza colagenului
În corpul uman, cele mai frecvente sunt următoarele: colagenul de tip I se găsește în principal în țesutul conjunctiv al pielii, tendoanelor, oaselor, corneei, sclerei și pereților arterelor; Colagenul de tip II face parte din cartilajul hialin și fibros, din corpul vitros și din cornee; Colagenul de tip III se găsește în dermul pielii fetale, în pereții vaselor mari de sânge și în fibrele reticulare ale organelor hematopoietice; Tipul IV - în membranele bazale, capsula cristalinului; Colagenul de tip V este prezent în corion, amnios, endomisium, perimisium, piele, în jurul celulelor (fibroblaste, endoteliale, mușchi neted) care sintetizează colagenul.
Moleculele de colagen au aproximativ 280 nm lungime și 1,4 nm lățime. Ele sunt construite din tripleți - trei lanțuri polipeptidice (lanțul α) ale precursorului de colagen - procolagen, răsucite într-o singură spirală în celulă. Acesta este primul molecular, nivelul de organizare a fibrei de colagen. Procolagenul este secretat în substanța intercelulară (Fig. 8.8).
Al doilea, supramolecular, nivel - organizarea extracelulară a fibrei de colagen - sunt molecule de tropocolagen agregate în lungime și reticulate prin legături de hidrogen, formate prin scindarea peptidelor procolagen terminale. În primul rând, se formează protofibrile, iar 5-6 protofibrile, legate între ele prin legături laterale, formează microfibrile cu o grosime de aproximativ 5 nm.
Cu participarea glicozaminoglicanilor, secretați și de fibroblaste, se formează al treilea, fibrilar, nivelul de organizare a fibrei de colagen. Fibrilele de colagen sunt striate în cruce

Orez. 8.9. Fibrilă de colagen (preparat de N. P. Omelyanenko). P - punct. Micrografie electronică. Crește cu 200.000
ny structuri cu o grosime medie de 20-100 nm. Perioada de repetiție a zonelor întunecate și luminoase este de 64-67 nm (Fig. 8.9, 8.10). Se crede că fiecare moleculă de colagen din rândurile paralele este compensată cu un sfert din lungime față de lanțul învecinat, provocând alternarea dungilor întunecate și deschise. În benzile întunecate sub un microscop electronic, sunt vizibile linii subțiri transversale secundare, cauzate de aranjarea aminoacizilor polari în moleculele de colagen.
Al patrulea, fibră, nivelul de organizare. Fibra de colagen, formată prin agregarea fibrilelor, are o grosime de 1-10 μm (în funcție de topografie). Include un număr variabil de fibrile - de la un singur la câteva zeci. Fibrele pot fi pliate în mănunchiuri de până la 150 microni grosime.
Fibrele de colagen se caracterizează prin alungire scăzută și rezistență ridicată la tracțiune. În apă, grosimea tendonului crește cu 50% ca urmare a umflăturilor, iar în acizi și alcali diluați - de 10 ori, dar fibra este scurtată cu 30%. Capacitatea de a se umfla este mai pronunțată la fibrele tinere. Când sunt tratate termic în apă, fibrele de colagen formează o substanță adezivă (greacă: colla- lipici), care a dat numele acestor fibre.
Tipuri de fibre de colagen sunt reticularȘi fibre precolagen. Acestea din urmă reprezintă forma inițială de formare a fibrei de colagen în embriogeneză și în timpul regenerării. Conțin colagen de tip III și o cantitate crescută de carbohidrați, care sunt sintetizați de celulele reticulare ale organelor hematopoietice. Ele formează o rețea tridimensională - reticulul, care le determină numele.
Fibre elastice. Prezența fibrelor elastice (fibra elasticae)în țesutul conjunctiv determină elasticitatea și extensibilitatea acestuia. Fibrele elastice sunt inferioare ca rezistență fibrelor de colagen. Forma secțiunii transversale a fibrelor este rotundă și aplatizată. În țesutul conjunctiv lax se anastomozează pe scară largă între ele. Grosimea fibrelor elastice este de obicei mai mică decât colagenul (0,2-1 microni), dar poate ajunge la câțiva micrometri (de exemplu, în ligamentul nucal). Compoziția fibrelor elastice se împarte în microfibrilareȘi amorf Componente.

Orez. 8.10. Formarea substanței intercelulare (după R. Krstic, cu modificări):
1 - fibroblast; 2 - lanțuri polipeptidice; 3 - molecule de tropocolagen; 4 - glicozaminoglicani; 5 - polimerizarea moleculelor de tropocolagen; 6 - protofibrila; 7 - fascicul de protofibrile (fibrila de colagen); 8 - molecula de elastina; 9 - fibrila elastica
Baza fibrelor elastice este glicoproteina globulară - elastina, sintetizat de fibroblaste și celule musculare netede (în primul rând, molecular, nivel de organizare). Elastină se caracterizează printr-un conținut ridicat de prolină și glicină și prin prezența a doi derivați ai aminoacizilor - desmozină și izodesmozină, care sunt implicați în stabilizarea structurii moleculare a elastinei și în conferirea acesteia elasticitate și elasticitate. Moleculele de elastină, având globule cu un diametru de 2,8 nm, în afara celulei sunt conectate în lanțuri - protofibrile de elastină cu o grosime de 3-3,5 nm (al doilea, supramolecular, nivel de organizare). Protofibrilele de elastină în combinație cu glicoproteina (fibrilina) formează microfibrile (Fig. 8.11) cu o grosime de 8-19 nm (al treilea, fibrilar, nivel de organizare). Al patrulea nivel de organizare este

Orez. 8.11. Structura ultramicroscopică a fibrei elastice: A- schema: 1 - piesa centrala omogena; 2 - microfibrile (conform lui Yu. I. Afanasyev); b- micrografie electronică, mărire 45.000 (pregătire de V. P. Slyusarchuk): 1 - partea centrală omogenă; 2 - microfibrile la periferia fibrei; 3 - complexul de fibroblast Golgi; 4 - reticul endoplasmatic; 5 - centriol
fibră. Cele mai mature fibre elastice conțin aproximativ 90% din componenta proteică elastică amorfă (elastină) în centru și microfibrile de-a lungul periferiei. În fibrele elastice, spre deosebire de fibrele de colagen, nu există structuri cu striații transversale pe lungimea lor.
Pe lângă fibre elastice mature, există elaunineȘi oxitalan fibre. În fibrele de elaunină, raportul dintre microfibrile și componenta amorfă este aproximativ egal, în timp ce fibrele oxitalane constau numai din microfibrile.
Componentă amorfă a substanței intercelulare. Celulele și fibrele țesutului conjunctiv sunt închise într-o componentă amorfă sau substanță fundamentală (substantia fundamentals). Această substanță asemănătoare gelului este un mediu multicomponent metabolic, tampon integrativ, care înconjoară structurile celulare și fibroase ale țesutului conjunctiv, nervilor și vaselor de sânge. Componentele substanței principale includ proteine din plasmă sanguină, apă, ioni anorganici, produse ale metabolismului celular, precursori solubili ai colagenului și elastinei, proteoglicanii, glicoproteinele și complexele formate de acestea. Toate aceste substanțe sunt în continuă mișcare și reînnoire (Fig. 8.12).
Proteoglicanii (PG) sunt compuși proteine-carbohidrați care conțin 90-95% carbohidrați.
Glicozaminoglicanii (GAG) sunt compuși polizaharidici care conțin de obicei acid hexuronic cu amino zaharuri (N-acetilglicozamină, acid N-acetilgalactic)
Proteoglican

Orez. 8.12. Organizarea moleculară a substanței țesutului conjunctiv amorf
tozamină). Moleculele GAG conțin multe grupări hidroxil, carboxil și sulfat care au o sarcină negativă; ele atașează cu ușurință moleculele de apă și ionii, în special Na + și, prin urmare, determină proprietățile hidrofile ale țesutului. GAG-urile sunt permeabile la oxigen și CO 2, dar protejează organele de pătrunderea corpurilor străine și a proteinelor. GAG-urile sunt implicate în formarea structurilor fibroase ale țesutului conjunctiv și proprietățile lor mecanice, procesele reparatorii ale țesutului conjunctiv și în reglarea creșterii și diferențierii celulelor. Dintre acești compuși, cel mai frecvent în tipuri de țesut conjunctiv este acidul hialuronic, precum și GAG-urile sulfatate: sulfati de condroitină (în cartilaj, piele, cornee), sulfat de dermatan (în piele, tendoane, în peretele vaselor de sânge etc. ), sulfat de keratan, sulfat de heparină (în compoziția multor membrane bazale).
Heparina este un GAG format din acid glucuronic și glicozamină. La oameni și animale, este produs de mastocite și este un factor anticoagulant natural în sânge.
Glicoproteinele (GP) sunt o clasă de compuși proteici cu oligozaharide (hexoze, hexoze, fucoze, acizi sialici), care fac parte atât din fibre, cât și din substanțe amorfe. Acestea includ GP solubile asociate cu proteoglicani; GP a țesuturilor calcificate; Medicii de familie legati de colagen. GP joacă un rol major în formarea structurii substanței intercelulare a țesutului conjunctiv și determină caracteristicile funcționale ale acestuia (fibronectină, condronectină, fibrilină, laminină etc.).
Fibronectina este principala suprafață GP a fibroblastelor. În spațiul intercelular este asociat în principal cu colagenul interstițial. Se crede că fibronectina determină lipiciitatea, motilitatea, creșterea și specializarea celulelor etc.
Fibrilina formează microfibrile și întărește legătura dintre componentele extracelulare.
Laminina este o componentă a membranei bazale, constând din trei lanțuri polipeptidice interconectate prin compuși disulfuri, precum și cu colagen de tip V și receptori de suprafață celulară.
Țesuturi conjunctive dense
Țesuturi conjunctive dense (textus connectivus compactus) caracterizat printr-un număr relativ mare de fibre situate dens și o cantitate mică de elemente celulare și principala substanță amorfă dintre ele. În funcție de natura locației structurilor fibroase, acestea se disting neorientat(neformat), orientat(formatat) și amestecat tipuri de țesut conjunctiv dens.
Neorientat un tip de țesut caracterizat printr-un aranjament dezordonat al fibrelor (de exemplu, derma pielii). ÎN orientat In functie de tipul de tesut, dispunerea fibrelor este strict ordonata si in fiecare caz corespunde conditiilor in care functioneaza organul (tendoane, ligamente, membrane fibroase). Amestecat tipul, de regulă, are o structură stratificată (cornee, sclera) cu direcții alternante ale elementelor fibroase de țesut conjunctiv dens.
Tendon(tendo). Este alcătuit din mănunchiuri paralele groase, strâns împachetate de fibre de colagen aranjate în paralel. Între aceste fascicule sunt situate fibrocite numite celule tendinoase (tendinocyti),și o cantitate mică de fibroblaste și material amorf măcinat. Procesele lamelare subțiri ale fibrocitelor intră în spațiile dintre fasciculele de fibre și sunt în contact strâns cu acestea. Fiecare mănunchi de fibre de colagen, separat de stratul adiacent de fibrocite, este numit un fascicul de ordinul întâi. Mai multe fascicule de ordinul întâi, înconjurate de straturi subțiri de țesut conjunctiv lax, alcătuiesc fasciculele de ordinul doi. Separă straturi de țesut conjunctiv lax grinzi de ordinul doi, numit endotenoniu. Din grinzi de ordinul doi sunt compuse grinzi de ordinul trei, separate de straturi mai groase de țesut conjunctiv lax - peritenoniu. Uneori, fasciculul de ordinul trei este tendonul în sine. Tendoanele mari pot avea, de asemenea, fascicule de ordinul al patrulea.
Peritenoniul și endotenoniul conțin vase de sânge care alimentează tendonul, nervii și terminațiile nervoase proprioceptive care trimit semnale către sistemul nervos central despre starea de tensiune în țesutul tendonului.
Ligamentul nucal aparține, de asemenea, tipului format de țesut conjunctiv dens, doar că este format din fibre elastice groase.
Unele tendoane de la atașamentele lor la oase sunt închise în teci formate din două teci de țesut conjunctiv fibros (vagin) intre care se afla un lichid (lubrifiant) bogat in acid hialuronic.
Membrane fibroase. Aceste structuri, construite din țesut conjunctiv dens, includ fascia, aponevroze, centrii tendinei diafragmei, capsule ale unor organe, dura mater, sclera, pericondriu, periost, precum și tunica albuginea a ovarului și a testiculului etc. Membranele fibroase ale acestui tip de țesut sunt greu de întins datorită faptului că fasciculele de fibre de colagen și fibroblastele și fibrocitele aflate între ele sunt dispuse într-o anumită ordine în mai multe straturi unul deasupra celuilalt. În fiecare strat, mănunchiuri ondulate de fibre de colagen se desfășoară paralel între ele într-o direcție, care nu coincide cu direcția în straturile adiacente (țesut conjunctiv organizat paralel în direcții diferite). Mănunchiuri individuale de fibre trec de la un strat la altul, conectându-le între ele. Pe lângă fasciculele de fibre de colagen, membranele fibroase conțin fibre elastice. Structurile fibroase precum periostul, sclera, tunica albuginea testiculară, capsulele articulare și altele se caracterizează printr-o aranjare mai puțin regulată a fasciculelor de fibre de colagen și un număr mare de fibre elastice în comparație cu aponevrozele.
8.1.2. Țesuturi conjunctive cu proprietăți deosebite
Astfel de țesături includ reticular, grasȘi membrană mucoasă Ele se caracterizează printr-o predominanță a celulelor omogene, cu care însăși denumirea acestor tipuri de țesut conjunctiv este asociată de obicei.
Țesut reticular
Țesut reticular (textus reticularis) este un tip de țesut conjunctiv, are o structură asemănătoare rețelei și constă în proces celule reticulareȘi fibre reticulare (argirofile).(Fig. 8.13). Majoritatea celulelor reticulare sunt asociate cu fibre reticulare și sunt conectate între ele prin procese, formând o rețea tridimensională. Țesutul reticular formează stroma organelor hematopoietice și micromediul pentru celulele sanguine care se dezvoltă în ele.
Fibrele reticulare (diametrul 0,5-2 µm) sunt un produs al sintezei celulelor reticulare. Ele sunt detectate la impregnarea cu săruri de argint, de aceea sunt numite și argirofile (din greacă. argentum- argint). Aceste fibre sunt rezistente la acizi slabi și alcalii și nu sunt digerate de tripsină.
În grupul fibrelor argirofile se face distincția între fibrele reticulare și cele precolagen. Fibrele reticulare în sine sunt formațiuni definitive, finale care conțin colagen de tip III. Fibrele reticulare, în comparație cu fibrele de colagen, conțin concentrații mari de sulf, lipide și carbohidrați. Într-un microscop electronic, fibrilele de fibre reticulare nu au întotdeauna striații clar definite cu o perioadă de 64-67 nm. În ceea ce privește extensibilitatea, aceste fibre ocupă
Au o pozitie intermediara intre colagen si elastic. Fibrele de precolagen reprezintă forma inițială de formare a fibrelor de colagen în embriogeneză și în timpul regenerării.

Orez. 8.13.Țesut reticular: A- micrografie a celulelor reticulare: 1 - nucleul unei celule reticulare; 2 - procese ale citoplasmei; b- microfotografie a fibrelor reticulare ale ganglionului limfatic: 1 - fibre reticulare. Impregnare cu azotat de argint
Țesut adipos
Țesut adipos (textul adipos)- acestea sunt acumulări de celule adipoase găsite în multe organe (vezi Adipocite). Există două tipuri de țesut adipos - albȘi maro. Acești termeni sunt condiționati și reflectă caracteristicile colorării celulelor. Țesutul adipos alb este larg răspândit în corpul uman, iar țesutul adipos maro se găsește în principal la nou-născuți și la unele animale (rozătoare și hibernatoare) de-a lungul vieții.
Țesut adipos alb la om este localizată sub piele, în special în partea inferioară a peretelui abdominal, pe fese și coapse, unde formează stratul adipos subcutanat, în epiploon, mezenter și regiunea retroperitoneală. Țesutul adipos este mai mult sau mai puțin clar împărțit de straturi de țesut conjunctiv fibros lax în lobuli de diferite dimensiuni și forme. Celulele adipoase din interiorul lobulilor sunt destul de apropiate unele de altele (Fig. 8.14). Fibroblastele, elementele limfoide și mastocitele sunt situate în spațiile înguste dintre ele. Fibrele subțiri de colagen sunt orientate între celulele adipoase în toate direcțiile. Capilarele sanguine și limfatice, situate în straturi de țesut conjunctiv fibros lax, înconjoară îndeaproape grupuri de celule adipoase sau lobuli de țesut adipos cu ansele lor. În țesutul adipos au loc procese active de metabolizare a acizilor grași, carbohidraților și formarea grăsimilor din carbohidrați.
În timpul descompunerii grăsimilor, se eliberează o cantitate mare de apă și se eliberează energie, astfel încât țesutul adipos joacă nu numai rolul unui depozit de substraturi pentru sinteza compușilor cu energie ridicată, ci indirect și rolul unui depozit de apă. .
În timpul postului, țesutul adipos subcutanat și perinefric, țesutul adipos omental și mezenteric își pierd rapid rezervele de grăsime. Picături de lipi
Grăsimile din interiorul celulelor sunt zdrobite, iar celulele adipoase capătă o formă stelata sau în formă de fus. În zona orbitală a ochilor și în pielea palmelor și tălpilor, țesutul adipos pierde doar o cantitate mică de lipide chiar și în timpul postului prelungit. Aici, țesutul adipos joacă un rol mai degrabă mecanic decât metabolic. În aceste locuri este împărțit în lobuli mici înconjurați de fibre de țesut conjunctiv.
Țesut adipos brunîntâlnit la nou-născuți și la unele animale pe gât, lângă omoplați, în spatele sternului, de-a lungul coloanei vertebrale, sub piele și între mușchi. Este format din celule adipoase dens împletite cu hemocapilare. Aceste celule iau parte la procesele de producere a căldurii. Adipocitele din țesutul adipos brun au multe incluziuni grase mici în citoplasmă (Fig. 8.15). În comparație cu celulele albe ale țesutului adipos, acestea au semnificativ mai multe mitocondrii. Culoarea brună a celulelor adipoase este dată de pigmenții care conțin fier – citocromii mitocondriali.
Capacitatea oxidativă a celulelor adipoase brune este de aproximativ 20 de ori mai mare decât a celulelor adipoase albe și de aproape 2 ori mai mare decât capacitatea oxidativă a mușchiului inimii. Când temperatura ambientală scade, activitatea proceselor oxidative din țesutul adipos brun crește. La

Orez. 8.14. Structura țesutului adipos alb (conform lui Yu. I. Afanasyev): A b- structura ultramicroscopică a adipocitelor: 1 - nucleul adipocitelor; 2 - picaturi mari de lipide; 3 - fibre nervoase; 4 - hemocapilare; 5 - mitocondriile adipocitelor; V- tesut adipos: 1 - adipocite; 2 - vas de sânge. Micrografie (colorarea Sudan III)

Orez. 8.15. Structura țesutului adipos maro (conform lui Yu. I. Afanasyev): A- adipocite cu grăsimea îndepărtată la microscop optic; b- structura ultramicroscopică a adipocitelor: 1 - nucleul adipocitelor; 2 - lipide zdrobite fin; 3 - numeroase mitocondrii; 4 - hemocapilare; 5 - fibre nervoase
Aceasta eliberează energie termică, care încălzește sângele din capilarele sanguine. În reglarea schimbului de căldură, un anumit rol îl joacă sistemul nervos simpatic și hormonii medulei suprarenale - adrenalina și norepinefrina, care stimulează activitatea lipazei tisulare, care descompune trigliceridele în glicerol și acid gras. Aceasta duce la eliberarea de energie termică, care încălzește sângele care curge în numeroasele capilare dintre adipocite. În timpul postului, țesutul adipos maro se modifică mai puțin decât țesutul adipos alb.
Țesut mucos
Țesut mucos (textus mucosus) apare în mod normal doar la nivelul embrionului. Obiectul clasic pentru studiul său este cordonul ombilical al fătului uman (Fig. 8.16). Elementele celulare de aici sunt reprezentate de un grup eterogen de celule care se diferențiază în mod divergent de celulele mezenchimale în perioada prenatală. Printre acestea, se disting fibroblastele mari în formă de stea, miofibroblastele și celulele musculare netede. Se disting prin capacitatea lor de a sintetiza vimentină, desmină, actină și miozină. Fibroblaste ale țesutului mucos al canalului ombilical -

Orez. 8.16.Țesut conjunctiv mucos din cordonul ombilical: 1 - mucocite; 2 - substanță intercelulară; 3 - peretele unui vas de sânge
tecul („jeleul lui Wharton”) sintetizează colagenul de tip IV, caracteristic membranelor bazale, lamininei și sulfatului de heparină.
În prima jumătate a sarcinii, acidul hialuronic se găsește în cantități mari între celulele acestui țesut, ceea ce determină consistența gelatinoasă a substanței principale. Fibroblastele țesutului conjunctiv gelatinos sintetizează slab proteine fibrilare. Numai în etapele ulterioare ale dezvoltării embrionului apar în substanța gelatinoasă fibrile de colagen aranjate vag.
8.2. ȚESUT SCHELETIC
Țesuturile scheletice (schelete de text)- sunt tipuri de tesut conjunctiv cu functie de sustinere, mecanica semnificativ pronuntata, datorita prezentei substantei intercelulare dense: cartilaj, tesut osos, dentina si ciment dentar. Pe lângă funcția lor principală, aceste țesuturi participă la metabolismul apă-sare.
8.2.1. Țesutul cartilajului
Țesutul cartilajului (textus cartilaginei) fac parte din organele sistemului respirator, articulațiile, discurile intervertebrale și altele, constau din celule - condrociteȘi condroblasteși o cantitate mare de substanță hidrofilă intercelulară, caracterizată prin elasticitate. Funcția de susținere a țesutului cartilajului este asociată cu aceasta. Țesutul proaspăt de cartilaj conține aproximativ 70-80% apă, 10-15% substanțe organice și 4-7% săruri. De la 50 la 70% din materia uscată a țesutului cartilajului este colagen. Țesutul cartilajului în sine nu are vase de sânge, iar nutrienții difuzează din țesutul înconjurător pericondriu.
Clasificare. Există trei tipuri de țesut cartilaj: hialină, elastică, fibroasă. Această diviziune a țesutului cartilajului se bazează pe caracteristicile structurale și funcționale ale structurii substanței lor intercelulare, gradul de conținut și raportul dintre colagen și fibre elastice.
Dezvoltarea țesutului cartilajului (condrogistogeneză)
Sursa dezvoltării țesutului cartilajului este mezenchimul. Inițial, în unele părți ale corpului embrionului, unde se formează cartilajul, celulele mezenchimale
ne pierdem procesele, ne înmulțim viguros și, strâns alăturați unul de celălalt, creăm o anumită tensiune - turgor. Se numesc astfel de zone rudimente condrogenice, sau insulițe condrogenice(Fig. 8.17). Conțin celule stem care se diferențiază în celule semi-stem condrogenice. Celulele stem se caracterizează printr-o formă rotundă, o valoare mare a raporturilor nuclear-citoplasmatice, un aranjament difuz al cromatinei și un nucleol mic. Organelele din aceste celule sunt slab dezvoltate. În celulele semi-stem (precondroblaste), numărul de ribozomi liberi crește, apar membrane reticulului endoplasmatic de tip granular, forma celulei se alungește, iar relațiile nuclearo-citoplasmatice scad. Ulterior, precursorii slab diferențiați se diferențiază în condroblaste identificabile morfologic (vezi Fig. 8.17).
Condroblaste(din greaca. chondros- cartilaj, blastos- rudiment) sunt celule tinere turtite capabile de proliferare si sinteza substantei intercelulare a cartilajului (proteoglicani). Citoplasma condroblastelor are un reticul endoplasmatic granular și agranular bine dezvoltat, complexul Golgi. Când este colorată, citoplasma condroblastelor se dovedește a fi bazofilă datorită conținutului bogat
Orez. 8.17. Dezvoltarea țesutului cartilaginos hialin (schemă conform lui Yu. I. Afanasyev): A- insuliță condrogenă; b- tesut cartilaginos primar; V- etapele diferenţierii condrocitelor. 1 - celule mezenchimale; 2 - celule cu diviziune mitotică; 3 - substanta intercelulara; 4 - substanță intercelulară oxifilă; 5 - substanță intercelulară bazofilă; 6 - condrocite tinere; 7 - hemocapilar

Orez. 8.18. Grupuri izogenice ale celulelor cartilajului:
1 - condrocit; 2 - matricea teritoriului celular, constând dintr-o rețea în buclă de proteine noncolagene și proteoglicani; 3 - fibre de colagen care formează peretele lacunei; 4 - secțiune interteritorială a cartilajului; 5 - proteoglicani în matricea interteritorială (după V.N. Pavlova, cu modificări)
ARN. Condroblastele sintetizează și secretă proteine fibrilare (colagen), apare o substanță intercelulară, care se colorează oxifil. Acesta este modul în care se formează țesutul primar de cartilaj. Cu participarea condroblastelor are loc creştere periferică (apoziţională). cartilaj. Odată cu diferențierea suplimentară a țesutului cartilajului, condroblastele se dezvoltă în condrocite.
Condrocite- principalul tip de celule ale țesutului cartilaginos. Au formă ovală, rotundă sau poligonală – în funcție de gradul de diferențiere. Sunt localizate în cavități speciale (lacune) din substanța intercelulară, individual sau în grup. Se numesc grupuri de celule situate într-o cavitate comună izogenic(din greaca. isos- egal, geneză- dezvoltare). Ele se formează prin diviziunea unei celule (Fig. 8.18). În grupele izogenice se disting trei tipuri de condrocite.
Primul tip de condrocite se caracterizează printr-un raport nuclear-citoplasmatic ridicat, dezvoltarea elementelor vacuolare ale complexului Golgi și prezența mitocondriilor și a ribozomilor liberi în citoplasmă. În aceste celule se observă adesea figuri de diviziune mitotică, ceea ce ne permite să le considerăm sursa apariției grupurilor izogenice de celule (Fig. 8.19). Condrocitele de tip I predomină la cartilajele tinere, în curs de dezvoltare. Condrocitele de tip II se caracterizează prin scăderea raportului nuclear-citoplasmatic, slăbirea sintezei ADN, păstrarea unui nivel ridicat de ARN, dezvoltarea intensivă a reticulului endoplasmatic granular și a tuturor componentelor complexului Golgi, care asigură formarea și secreția de glicozaminoglicanii și proteoglicanii în substanța intercelulară. Condrocitele de tip III se caracterizează prin raportul nuclear-citoplasmatic cel mai scăzut, dezvoltarea puternică și aranjarea ordonată a reticulului endoplasmatic granular. Aceste celule își păstrează capacitatea de a forma și secreta proteine, dar sinteza lor de glicozaminoglicani este redusă.
Pe măsură ce anlagul cartilaginos se dezvoltă la periferie la granița cu mezenchimul, pericondriu- o înveliș care acoperă cartilajul în curs de dezvoltare din exterior și constând dintr-un strat exterior fibros și intern condrogen (cambial). În zona condrogenă, celulele se divid și se diferențiază intens în condroblaste, care își păstrează capacitatea de a sintetiza ADN-ul, de a se reproduce și, de asemenea, de a sintetiza componente ale substanței intercelulare (colagenul de tip I și III). În procesul de secreție a produselor de sinteză și stratificarea pe cartilajul existent de-a lungul periferiei sale, ei înșiși
celulele sunt „împodobite” în produsele activității lor. Așa crește cartilajul prin suprapunere sau creştere apoziţională.
Celulele cartilajului aflate în centrul cartilajului tânăr în curs de dezvoltare își păstrează capacitatea de a se diviza mitotic pentru o perioadă de timp, rămânând în aceeași lacună (grupuri izogenice de celule) și produc colagen de tip II. Datorită creșterii numărului acestor celule și producției lor de substanță intercelulară, masa cartilajului crește din interior, ceea ce se numește creșterea interstițială. Creșterea interstițială se observă în embriogeneză, precum și în timpul regenerării țesutului cartilajului.
Odată cu creșterea și dezvoltarea cartilajului, secțiunile sale centrale se îndepărtează din ce în ce mai mult de vasele din apropiere și încep să întâmpine dificultăți în nutriție, efectuate difuz din vasele pericondrului. Ca urmare, condrocitele își pierd capacitatea de a se reproduce, unele dintre ele sunt distruse, iar proteoglicanii sunt transformați într-o proteină oxifilă mai simplă - albumoid.
Astfel, diferenţialul condroblastic în histogeneza embrionară este reprezentat de toate formele de celule în maturare. Cu toate acestea, ulterior include doar legăturile mijlocii și finale (condroblaste și condrocite) și este singura din țesuturile cartilaginoase.

Orez. 8.19. Trei tipuri de condrocite (conform lui Yu. I. Afanasyev):
A- Tipul I; b - Tipul II; V - tipul III. 1 - celulă care se divide mitotic; 2 - reticul endoplasmatic; 3 - mitocondrii; 4 - glicogen; 5 - substanta intercelulara
Țesut cartilaj hialin
Țesut cartilaj hialin (textus cartilagineus hyalinus), numit și vitros (din greacă. hialos- sticlă) - datorită transparenței și culorii alb-albăstrui, este cea mai comună varietate -

Orez. 8.20. Cartilajul hialin: A- microfotografia cartilajului hialin al traheei: 1 - pericondriu; 2 - condrocite tinere; 3 - substanță de bază cu grupări izogenice de condrocite situate în interiorul acesteia (4)
densitatea țesutului cartilajului. În corpul adult, țesutul hialin se găsește la joncțiunea coastelor cu sternul, în laringe, căile respiratorii și pe suprafețele articulare ale oaselor.
Țesutul cartilajului hialin al diferitelor organe are multe în comun, dar în același timp diferă în specificul organului - locația celulelor, structura substanței intercelulare. Majoritatea țesutului cartilajului hialin găsit în corpul uman este acoperit pericondriuși reprezintă formațiuni anatomice - cartilaj.
Pericondrul are două straturi: stratul exterior, format din țesut conjunctiv fibros cu vase de sânge; interne, predominant celulare, conținând condroblaste și precursorii acestora – precondroblastele. Sub pericondriu în stratul superficial există condrocite tinere în formă de fus, a căror axă lungă este îndreptată de-a lungul suprafeței cartilajului (Fig. 8.20). În straturile mai profunde, celulele cartilajului capătă o formă ovală sau rotundă. Datorită faptului că procesele sintetice și secretoare din aceste celule sunt slăbite, după divizare ele nu diverg departe, ci zac compact, formând așa-numitele grupări izogenice de 2-4 condrocite.
Celulele cartilajului mai diferențiate și grupările izogenice, în plus față de stratul pericelular oxifil al substanței intercelulare, au o zonă bazofilă situată în exterior. Aceste proprietăți se explică prin distribuția neuniformă a componentelor chimice ale substanței intercelulare - proteine și glicozaminoglicani.
În cartilajul hialin de orice localizare se obișnuiește să se distingă zone teritoriale substanță intercelulară sau matrice (vezi Fig. 8.18). Regiunea teritorială include matricea care înconjoară imediat celulele cartilajului sau grupurile acestora. În aceste zone, fibrele și fibrilele de colagen de tip II, răsucindu-se, înconjoară grupuri izogenice de celule de cartilaj, protejându-le de presiunea mecanică. În interteritorial

Orez. 8.20. Continuare
b- diagrama structurii cartilajului și pericondrului (Fig. Yu. I. Afanasyev): 1 - strat fibros exterior; 2 - stratul celular interior; 3 - țesut cartilaginos; V- componente celulare și fibroase ale cartilajului articular (conform lui V.P. Modyaev, V.N. Pavlova, cu modificări). I - zona de suprafata; II - zona intermediara; III - zona bazală (profundă); IV - os subcondral; A - componentele celulare ale cartilajului articular: 1 - placa acelulară; 2 - condrocite ale stratului tangențial; 3 - condrocite ale zonei de tranziție; 4 - grupări izogenice; 5 - „coloane” de condrocite; 6 - condrocite hipertrofiate; 7 - linia bazofilă (de frontieră) între cartilajul calcificat și cel necalcificat; 8 - cartilaj calcifiant; B - sistemul fibrilar al cartilajului articular: 1 - placa acelulară; 2 - fibre tangențiale ale zonei de suprafață; 3 - direcțiile principale ale fibrelor de colagen în zona intermediară; 4 - fibre radiale ale stratului bazal; 5 - linie bazofilă (limită).
În această matrice, fibrele de colagen sunt orientate în direcția vectorului de acțiune a forțelor sarcinilor principale. Spațiul dintre structurile de colagen este umplut cu proteoglicani.
Condronectina joacă un rol important în organizarea structurală a substanței intercelulare a cartilajului. Această glicoproteină conectează celulele între ele și la diferite substraturi (colagen, glicozaminoglicani). Funcția biomecanică de susținere a țesutului cartilajului în timpul compresiei și tensiunii este asigurată nu numai de structura cadrului fibros al acestuia, ci și de prezența proteoglicanilor hidrofili cu un nivel ridicat de hidratare (65-85%). Hidrofilitatea ridicată a substanței intercelulare favorizează difuzia nutrienților și a sărurilor. Gazele și mulți metaboliți difuzează, de asemenea, liber prin el. Cu toate acestea, moleculele mari de proteine cu proprietăți antigenice nu trec. Așa se explică transplantul de succes al secțiunilor de cartilaj în clinică (transplant de la o persoană la alta). Metabolismul condrocitelor este predominant anaerob, glicolitic.
Cu toate acestea, nu toate cartilajele sunt construite la fel. O caracteristică structurală a cartilajului hialin al suprafeței articulare este absența pericondrului pe suprafața orientată spre cavitatea articulară. Cartilajul articular este format din trei zone slab delimitate: superficial, intermediar și bazal (vezi Fig. 8.20, c).
În zona superficială a cartilajului articular există condrocite mici, turtite, nespecializate, care amintesc de fibrocite în structură.
În zona intermediară, celulele sunt mai mari, de formă rotundă, foarte active din punct de vedere metabolic: cu mitocondrii mari, un reticul endoplasmatic granular bine dezvoltat și un complex Golgi cu numeroase vezicule.
Zona profundă (bazală) este împărțită de o linie bazofilă în straturi necalcificante și calcifiante. Acesta din urmă este pătruns de vasele de sânge din osul subcondral dedesubt. O caracteristică a substanței intercelulare din zona profundă a cartilajului articular este conținutul de vezicule cu matrice densă - structuri membranare cu un diametru de 30 nm până la 1 μm, care sunt locurile de mineralizare inițială a țesuturilor scheletice (pe lângă cartilaj, acestea se găsesc în țesutul osos și predentină). Structurile membranare sunt formate prin bombarea unei secțiuni a plasmalemei condrocitelor (respectiv, osteoblast în țesutul osos și odontoblast în predentină), urmat de înmugurire de la suprafața celulei și distribuție localizată în zonele de mineralizare. Ele pot fi, de asemenea, un produs al dezintegrarii complete a celulelor. Cartilajul articular este hrănit doar parțial din vasele zonei profunde, dar în principal din lichidul sinovial al cavității articulare.
Țesut cartilaj elastic
Țesut cartilaj elastic (textus cartilagineus elasticus) se găsește în acele organe în care baza cartilaginoasă este supusă îndoirii (în auriculă, cartilajele corniculate și în formă de pană ale laringelui etc.). Într-o stare proaspătă, nefixată, țesutul cartilaginos elastic este de culoare gălbuie și nu la fel de transparent ca țesutul hialin. Conform structurii generale a cartilajului elastic, acesta este similar cu cartilajul hialin. La exterior este acoperit cu pericondriu. Celulele cartilajului (tinere și specializate condrocite) sunt localizate individual în capsule sau formează grupări izogenice. Una dintre principalele diferențe
O caracteristică semnificativă a cartilajului elastic este prezența în substanța sa intercelulară, alături de fibrele de colagen, a fibrelor elastice care pătrund în substanța intercelulară în toate direcțiile (Fig. 8.21).
Din stratul adiacent pericondului, fibrele elastice trec fără întrerupere în fibrele elastice ale pericondrului. Există mai puține lipide, glicogen și sulfați de condroitină în cartilajul elastic decât în cartilajul hialin.

Orez. 8.21. Cartilaj elastic. Micrografie, colorare - orceină: 1 - grupări izogenice ale condrocitelor; 2 - fibre elastice
Țesut cartilaj fibros
Țesut cartilaj fibros (textus cartilagineus fibrosa) situat în discurile intervertebrale, articulațiile semimobile, în locurile de tranziție a țesutului conjunctiv fibros (tendoane, ligamente) în cartilaj hialin, unde mișcările limitate sunt însoțite de tensiuni puternice. Substanța intercelulară conține mănunchiuri paralele de colagen,
afânându-se treptat și transformându-se în cartilaj hialin. Cartilajul are cavități în care sunt închise celulele cartilajului. Acestea din urmă sunt localizate individual sau formează mici grupări izogenice. Citoplasma celulelor este adesea vacuolată. Pe măsură ce cartilajul hialin se mișcă spre tendon, fibrocartilajul devine din ce în ce mai asemănător tendonului. La marginea cartilajului și tendonului, între fasciculele de colagen, se află coloane de celule cartilaginoase comprimate, care, fără nicio margine, trec în celulele tendinoase situate în țesutul conjunctiv dens (Fig. 8.22).
Modificări de vârstă. Pe măsură ce corpul îmbătrânește, concentrația de proteoglicani în țesutul cartilajului și hidrofilitatea asociată scad. Procesele de reproducere a condroblastelor și a condrocitelor tinere sunt slăbite. În citoplasma acestor celule, volumul complexului Golgi, reticulul endoplasmatic granular și mitocondriile scade, iar activitatea enzimatică scade.
Participa la resorbția celulelor modificate distrofic și a substanței intercelulare condroclaste, morfologic identice cu osteoclastele. După moartea condrocitelor, unele dintre goluri sunt umplute cu substanță amorfă și fibrile de colagen. În locurile intercelulare

Orez. 8.22.Țesut cartilaj fibros. Sectiunea discului intervertebral: 1 - fibre de colagen; 2 - condrocite
În substanță sunt detectate depozite de săruri de calciu („cretarea cartilajului”), în urma cărora cartilajul devine tulbure, dur și casant. Ca urmare, perturbarea rezultată a trofismului zonelor centrale ale cartilajului poate duce la creșterea în interior a vaselor de sânge în ele, urmată de formarea osului.
Regenerare. Regenerarea fiziologică a țesutului cartilajului se realizează datorită celulelor slab diferențiate ale pericondului și cartilajului, prin reproducerea și diferențierea precondroblastelor și condroblastelor. Cu toate acestea, acest proces este foarte lent. Regenerarea post-traumatică a țesutului cartilajului de localizare extraarticulară se realizează datorită pericondrului (Fig. 8.23).
În cartilajul articular, în funcție de adâncimea leziunii, regenerarea are loc atât datorită proliferării celulelor numai în grupe izogenice (cu afectare superficială), cât și datorită celei de-a doua surse de regenerare - celulele cambiale ale țesutului osos subcondral (cu afectare profundă a cartilaj), care formează matricea organică a osului - osteoid.
În orice caz, procesele distrofice (necrotice) sunt observate direct în zona leziunii țesutului cartilajului, iar condroblastele proliferante sunt localizate mai centrifug. În rană se formează țesut conjunctiv fibros, care este ulterior înlocuit cu cartilaj. În general, regenerarea este finalizată la 3-6 luni după accidentare.
Factori care reglează metabolismul țesutului cartilajului. Reglarea metabolismului țesutului cartilajului are loc sub influența sarcinii mecanice, a factorilor nervoși și hormonali. Presiunea periodică asupra țesutului cartilajului și slăbirea încărcăturii sunt factori care acționează constant pentru difuzarea nutrienților dizolvați în apă, a produselor metabolice și a regulatorilor hormono-umorali din capilarele pericondului, care are receptori și efectori, sau lichidul sinovial al articulațiilor. . În plus, condrocitele au citoreceptori pentru o serie de hormoni care circulă în sânge. Deci, hormonii hipofizari sunt somatotropina
și prolactina - stimulează creșterea țesutului cartilajului, dar nu afectează maturizarea acestora. Hormonii tiroidieni - tiroxina și triiodotironina - accelerează citodiferențierea condrocitelor, dar inhibă procesele de creștere în cartilaj. Hormonii tiroidei și ai glandelor paratiroide - calcitonina și hormonul paratiroidian - au un efect similar asupra metabolismului cartilajului, ajută la stimularea proceselor de creștere, dar într-o măsură mai mică la maturizarea acestora. Hormonul insulelor endocrine ale pancreasului - insulina - intensifică citodiferențierea celulelor mezenchimice scheletogene, iar în etapele ontogenezei postnatale are un efect de creștere și mitogen. Hormonii cortexului suprarenal - glucocorticoizii și hormonul sexual feminin estrogen - inhibă biosinteza colagenului și a glicozaminoglicanilor în condrocite, iar în perioada postnatală timpurie concentrațiile mari ale acestora contribuie la îmbătrânirea țesutului cartilajului și la modificările distructive ale acestuia. Hormonul sexual masculin - testosteronul - stimulează biosinteza glicozaminoglicanilor nesulfatați, ceea ce duce la scăderea maturării țesutului cartilajului. În general, trebuie remarcat faptul că hormonii reglează procesele metabolice specifice în condrocite, dar capacitatea condrocitelor de a răspunde la acțiunea lor depinde atât de starea stării endocrine a organismului (normal, deficiență sau exces de hormoni), cât și de starea structurală și funcțională a condrocitelor în sine.
8.2.2. Țesut osos
Țesut osos (textus ossei) este un tip polidiferent de țesut conjunctiv specializat cu mineralizare ridicată a intercelularului

Orez. 8.23. Regenerarea post-traumatică a cartilajului hialin cu puțin adâncime (a 1Și
b 1 şi adânc (a 2și b 2) daune: A\ 2 - regenerarea cartilajului articular: 1 - membrana sinoviala; 2 - cartilaj; 3 - os; 4 - zona de necroză; 5 - zona de proliferare; 6 - cartilaj necalcificat; 7 - cartilaj calcificat; 8 - osteoni cu vase; 9 - măduvă osoasă; 10 - tesut de granulatie; b - regenerarea cartilajului costal: 1 - pericondriu; 2 - cartilaj; 3 - zona de necroză; 4 - zona de proliferare; 5 - țesut de granulație (după V.N. Pavlova). Săgeți - direcțiile de mișcare ale celulelor și țesuturilor în procesul plăgii
materie organică care conține aproximativ 70% compuși anorganici, în principal fosfați de calciu. Peste 30 de oligoelemente (cupru, stronțiu, zinc, bariu, magneziu etc.) au fost găsite în țesutul osos, jucând un rol vital în procesele metabolice din organism.
Materie organică - matrice tesutul osos – reprezentat in principal de proteine si lipide de tip colagen. În comparație cu matricea țesutului cartilajului, acesta conține o cantitate relativ mică de apă, acid condroitinsulfuric, dar mulți acizi citric și alți acizi care formează complexe cu calciu, impregnând matricea organică a osului. Componentele organice și anorganice în combinație între ele determină proprietățile mecanice - capacitatea de a rezista la întindere, compresie etc. Dintre toate tipurile de țesuturi conjunctive, funcțiile de susținere, mecanice și de protecție sunt cele mai pronunțate în țesutul osos. Pentru organele interne, este și un depozit de săruri de calciu, fosfor etc.
În ciuda gradului ridicat de mineralizare, țesuturile osoase suferă o reînnoire constantă a substanțelor lor constitutive, distrugere și creare constantă și adaptare datorită condițiilor de operare în schimbare. Proprietățile morfofuncționale ale țesutului osos se modifică în funcție de vârstă, activitatea musculară, condițiile nutriționale, precum și sub influența activității glandelor endocrine, inervație etc.
Clasificare. Există două tipuri principale de țesut osos: fibros grosier (fibros cu plasă)Și lamelar. Aceste tipuri de țesut osos diferă ca structură și proprietăți fizice, care sunt determinate în principal de structura substanței intercelulare. Tesuturile osoase includ, de asemenea dentinăȘi ciment dinții asemănători țesutului osos în ceea ce privește gradul ridicat de mineralizare a substanței intercelulare (vezi capitolul 16) și funcția mecanică de susținere.
Țesut osos fibros dur
Țesut osos fibros dur (textus osseus reticulofibrosus) apare mai ales la embrioni. La adulți, poate fi găsit la locul suturilor craniene supra-crescute, la locurile de atașare a tendoanelor de oase. Fibrele de colagen dispuse aleatoriu formează mănunchiuri groase în el, clar vizibile chiar și la măriri relativ mici ale microscopului (Fig. 8.24).
În substanța principală a țesutului osos cu fibre grosiere există forme ovale alungite cavități osoase, sau lacune, cu tubuli lungi anastomozați în care se află celulele osoase - osteocitelor cu lăstarii lor. Suprafața osului este acoperită periost.
Țesut osos lamelar
Țesut osos lamelar (textus osseus lamellaris)- cel mai frecvent tip de tesut osos in corpul adultului. Se compune din plăci osoase (lamele ossea). Grosimea și lungimea acestuia din urmă

Orez. 8.24. Structura țesutului osos fibros grosier (conform lui Yu. I. Afanasyev): 1 - mănunchiuri de fibre de colagen care se împletesc; 2 - osteocite
variază de la câteva zeci la sute de micrometri, respectiv. Nu sunt monolitice, dar conțin fibrile de colagen (oseină) direcționate paralel, orientate în planuri diferite. În partea centrală a plăcilor, fibrilele au o direcție predominant longitudinală; de-a lungul periferiei se adaugă direcțiile tangențiale și transversale. Laminele se pot delamina, iar fibrilele unei lamine pot continua în cele adiacente, creând o singură bază fibroasă a osului. În plus, plăcile osoase sunt pătrunse cu fibrile și fibre individuale orientate perpendicular pe plăcile osoase, țesute în straturile intermediare dintre ele, datorită cărora se obține o rezistență mai mare a țesutului osos lamelar (Fig. 8.25). Substanța compactă și spongioasă din majoritatea oaselor plate și tubulare ale scheletului este construită din acest țesut.
Dezvoltarea țesutului osos (osteohistogeneză)
Dezvoltarea țesutului osos în embrion se realizează în două moduri: 1) direct din mezenchim (osteogeneză directă); 2) din mezenchim în locul unui model de os cartilaginos dezvoltat anterior (osteogeneză indirectă). Dezvoltarea postembrionară a țesutului osos are loc în timpul regenerării fiziologice și reparatorii.
Tesutul osos include osteoblasticȘi diferențiale osteoclastice. Primul (principal) constă dintr-un număr de celule care se diferențiază: celule stem, celule semi-stem (preosteoblaste), osteoblaste (un tip

Orez. 8.25. Structura țesutului osos lamelar (conform lui Yu. I. Afanasyev):
1 - plăci osoase; 2 - osteocite; 3 - contacte ale proceselor osteocitare; 4 -
fibre de colagen orientate în interiorul fiecărei plăci osoase
paralel
fibroblaste), osteocite. Procesele de diferențiere osteogenică a celulelor sunt influențate de factori osteogeni (proteina morfogenetică osoasă), presiunea parțială a oxigenului în țesut, prezența fosfatazei alcaline etc. Al doilea diferențial (asociat) include osteoclaste(un tip de macrofag) care se dezvoltă din celulele stem din sânge.
Celulele stem și semi-stem osteogene nu sunt identificate morfologic.
Osteoblastele(din greaca. osteon- os, blastos- rudiment) sunt celule tinere care creează țesut osos. În osul format, ele se găsesc numai în straturile profunde ale periostului și în locurile de regenerare a țesutului osos după leziune. Sunt capabili de proliferare, în osul care se formează acopera întreaga suprafață a fasciculului osos în curs de dezvoltare cu un strat aproape continuu (Fig. 8.26). Forma osteoblastelor poate fi diferită: cubică, piramidală sau unghiulară. Dimensiunea corpului lor este de aproximativ 15-20 de microni. Nucleul are formă rotundă sau ovală, adesea situat excentric și conține unul sau mai mulți nucleoli.

Orez. 8.26. Osteogeneza „directă”:
A- insula osteogenă (diagrama); b- stadiul (schema) osteoid; V- osificarea substantei intercelulare (schema); G- osteogeneză „directă” în osul plat (micrograf). 1 - celule mezenchimale; 2 - capilare sanguine; 3 - osteoblaste; 4 - osteoid; 5 - substanță intercelulară mineralizată; 6 - osteocit; 7 - osteoclaste
În citoplasma osteoblastelor, reticulul endoplasmatic granular, mitocondriile și complexul Golgi sunt bine dezvoltate (Fig. 8.27). În el sunt detectate cantități semnificative de ARN și activitate ridicată a fosfatazei alcaline. Osteoblastele secretă vezicule matricei care conțin lipide,

Orez. 8.27. Structura unui osteoblast (conform lui Yu. I. Afanasyev):
a - la nivel luminos-optic; b- la nivel ultramicroscopic. 1 - miez; 2 - citoplasmă; 3 - reticul endoplasmatic granular; 4 - osteoid; 5 - tesut osos mineralizat
Ca 2+, fosfatază alcalină, care duce la calcificarea matricei tisulare organice.
Osteocite(din greaca. osteon- os, cytus- celula) este numărul predominant de celule definitive ale țesutului osos care și-au pierdut capacitatea de a se diviza. Au o formă ramificată, un nucleu compact, relativ mare și citoplasmă ușor bazofilă (Fig. 8.28). Organelele sunt slab dezvoltate. Prezența centriolilor în osteocite nu a fost stabilită.
Celulele osoase se află în cavitățile osoase, sau lacune, care urmează contururile osteocitelor. Lungimea cavităților variază de la 22 la 55 de microni, lățimea - de la 6 la 14 microni. Tubulii cavităților osoase sunt umpluți cu lichid tisular, se anastomozează unul cu celălalt și cu peri-
spațiile sculare ale vaselor care intră în os. Schimbul de substanțe între osteocite și sânge are loc prin lichidul tisular.
Osteoclaste(din greaca. osteon- os și clastos- fragmentate). Aceste celule de natură hematogenă sunt capabile să distrugă cartilajul și oasele calcificate. Diametrul lor ajunge la 150-180 microni, conțin de la 3 la câteva zeci de nuclee (Fig. 8.29). Citoplasma este ușor bazofilă, uneori oxifilă. Osteoclastele sunt de obicei localizate pe suprafața trabeculelor osoase. Pe partea osteoclastei care este adiacentă suprafeței distruse, există o margine micropliată (onduită); este zona de sinteză și secreție a enzimelor hidrolitice. De-a lungul periferiei osteoclastei există o zonă de aderență strânsă a celulei la suprafața osoasă, care, parcă, sigilează zona de acțiune a enzimelor. Această zonă a citoplasmei este ușoară și conține puține organite, cu excepția microfilamentelor formate din actină.
Stratul periferic de citoplasmă de deasupra marginii ondulate conține numeroase vezicule mici și vacuole mai mari. Se crede că osteoclastele eliberează CO 2 în mediu, iar enzima carbonan hidrază, găsită aici, favorizează formarea acidului (H 2 CO 3) și dizolvarea compușilor de calciu. Osteoclastul este bogat în mitocondrii și lizozomi, ale căror enzime (colagenaza și alte proteaze) descompun colagenul și proteoglicanii din matricea țesutului osos. În locul în care osteoclastul intră în contact cu substanța osoasă, în aceasta din urmă se formează o lacună de resorbție. Un osteoclast poate distruge la fel de mult os cât creează 100 de osteoblaste în același timp. Funcțiile osteoblastelor
STS și osteoclaste sunt interconectate și corelează cu participarea hormonilor, prostaglandinelor, încărcăturii funcționale, vitaminelor etc.
Substanță intercelulară (substanța intercellularis) constă dintr-o substanță amorfă bazică impregnată cu săruri anorganice, în care se află fibrele de colagen, formând mănunchiuri mici. Conțin (până la 90%) proteine - colagen de tip I. Fibrele pot avea o direcție aleatorie (în țesutul osos grosier-fibros) sau strict orientată (în țesutul osos lamelar).
În comparație cu cartilajul, substanța de bază a țesutului osos conține o cantitate relativ mică de acid condroitinsulfuric, dar o mulțime de acizi citric și alți acizi care formează complexe cu calciul și impregnează matricea organică a osului. Pe lângă proteina de colagen, în substanța principală a țesutului osos se găsesc proteine non-colagen (osteocalcină, sialoproteină, osteonectină, osteopontin etc., implicate în procesele de mineralizare), precum și glicozaminoglicanii. Substanța fundamentală a osului conține cristale de hidroxiapatită dispuse în mod ordonat în raport cu fibrilele matricei organice a osului, precum și fosfat de calciu amorf. În țesutul osos au fost găsite peste 30 de microelemente (cupru, stronțiu, zinc, bariu, magneziu etc.), care joacă un rol important în procesele metabolice din organism. O creștere sistematică a activității fizice duce la o creștere a masei osoase de la 10 la 50% datorită mineralizării ridicate.
Osteohistogeneza directă. Această metodă de osteogeneză este tipică pentru dezvoltarea țesutului osos fibros grosier în timpul formării platului
Orez. 8.28. Structura unui osteocite (conform lui Yu. I. Afanasyev):
A- la nivel luminos-optic; b- la nivel ultramicroscopic. 1 - procesele osteocitelor; 2 - miez; 3 - reticul endoplasmatic; 4 - complexul Golgi; 5 - mitocondrii; 6 - substanță osoasă osteoidă (necalcificată) de-a lungul marginii golului, în care sunt situate osteocitele

Orez. 8.29. Structura unui osteoclast (conform lui Yu. I. Afanasyev):
A- la nivel luminos-optic; b- la nivel ultramicroscopic. 1 - miez; 2 - marginea ondulată a osteoclastei; 3 - zona de lumina; 4 - lizozomi; 5 - zona de resorbție a substanței intercelulare; 6 - substanta mineralizata
oase, cum ar fi oasele tegumentare ale craniului. Acest proces se observă în principal în prima lună de dezvoltare intrauterină și se caracterizează prin formarea de țesut osos osteoid primar „pântuit”, urmată de impregnarea (depunerea) de calciu, fosfor și alte săruri în substanța intercelulară. La prima etapă- formarea unei insulițe scheletice - în locurile de dezvoltare ale viitorului os are loc reproducerea focală a celulelor mezenchimale și vascularizarea insulei scheletice. Celulele mezenchimice se orientează în funcție de vectorii de încărcare și se diferențiază în progenitori osteogeni - preosteoblasti. La a doua etapă celulele insulare se diferențiază în osteoblaste, în care începe biosinteza proteinelor de colagen și secreția acesteia, în urma căreia apare o substanță intercelulară oxifilă cu fibrile de colagen - matricea organică a țesutului osos (stadiul osteoid). Fibrele în creștere împing celulele în afară, care rămân conectate între ele cu ajutorul proceselor de dezvoltare. Astfel, osteoblastele capătă o formă de proces și devin osteocite incluse în grosimea masei fibroase și își pierd capacitatea de a se reproduce. Mucoproteinele (osteomucoide) apar în substanța principală, cimentând fibrele într-o singură masă puternică. În același timp, din mezenchimul din jur se formează noi generații de osteoblaste, care cresc osul din exterior (creștere apozițională).
A treia etapă- calcificarea (impregnarea cu săruri) a substanţei intercelulare. În acest caz, osteoblastele secretă enzima fosfataza alcalină, care descompune glicerofosfații conținuti în sângele periferic în compuși carbohidrați (zaharuri) și acid fosforic. Acesta din urmă reacționează cu sărurile de calciu, care se depun în substanța și fibrele principale, mai întâi sub formă de compuși de calciu care formează amorfi.
Din aceasta se formează depozite de [Ca 3 (PO 4) 2], ulterior cristale de hidroxiapatită [Ca 10 (PO 4) 6 (OH) 2].
Unul dintre mediatorii calcificării este osteonectină- o glicoproteină care leagă selectiv sărurile de calciu și fosfor de colagen. Ca rezultat al calcificării, se formează bare transversale osoase sau grinzi. Apoi excrescențele se ramifică din aceste bare transversale, interconectându-se și formând o rețea largă. Spațiile dintre barele transversale se dovedesc a fi ocupate de țesut conjunctiv fibros lax, prin care trec vase de sânge.
Până la finalizarea histogenezei, un număr mare de fibre și celule osteogene apar în țesutul conjunctiv embrionar de-a lungul periferiei germenului osos. O parte din acest țesut conjunctiv fibros adiacent direct la trabeculele osoase se dezvoltă în periost, care asigură trofismul și regenerarea osoasă. Un astfel de os, care apare în stadiile dezvoltării embrionare și constă din lupte din țesut osos grosier fibros, se numește os spongios primar. În stadiile ulterioare de dezvoltare, este înlocuit cu osul spongios secundar al adulților, care diferă de primul prin faptul că este construit din țesut osos lamelar (a patra etapă a osteogenezei). Dezvoltarea țesutului osos lamelar este strâns legată de procesul de distrugere a secțiunilor individuale de os și de creșterea în interior a vaselor de sânge în grosimea osului cu fibre grosiere. Osteoclastele iau parte la acest proces atât în timpul osteogenezei embrionare, cât și după naștere (vezi Fig. 8.29). În jurul vaselor de sânge se formează un strat de osteoblaste și ulterior apar noi plăci de osteon concentrice. Fibrele de colagen din fiecare lamină sunt paralele, dar orientate în unghi față de fibrele laminei precedente. Astfel, în jurul vasului se formează cilindri osoși, introduși unul în celălalt (osteoni primari). Din momentul în care apar osteonii, țesutul osos fibros grosier încetează să se dezvolte și este înlocuit cu țesut osos lamelar. Din partea laterală a periostului se formează plăci înconjurătoare externe (generale, generale), care acoperă întregul os din exterior. Așa se dezvoltă oasele plate. Ulterior, osul format în perioada embrionară suferă o restructurare: osteonii primari sunt distruși și se dezvoltă noi generații de osteoni. Această restructurare a osului durează practic toată viața.
Spre deosebire de țesutul cartilajului, osul crește întotdeauna prin aplicarea de țesut nou pe țesutul existent, adică prin apoziții, iar diferențierea celulelor insulare scheletogene necesită o alimentare optimă cu sânge.
Osteohistogeneza indirectă. La luna a 2-a de dezvoltare embrionară, în locurile viitoarelor oase tubulare, din mezenchim se formează un rudiment cartilaginos care ia foarte repede forma viitorului os (model cartilaginos). Rudimentul este format din cartilaj hialin embrionar acoperit de pericondriu (Fig. 8.30). De ceva timp crește atât datorită celulelor formate pe partea laterală a pericondului, cât și datorită proliferării celulelor în zonele interne.

Orez. 8.30. Osteogeneză indirectă (cartilaginoasă). Formarea unui model cartilaginos de os și manșetă osului pericondral (conform lui Yu. I. Afanasyev):
a-d - stadiile osteogenezei. 1 - model cartilaginos primar al unui os tubular; 2 - pericondriu; 3 - țesut cartilaginos; 4 - manșetă os pericondral; 5 - periost; 6 - coloane de celule de cartilaj; 7 - zona celulelor veziculare; 8 - mezenchim care crește în cartilaj cu osteoclaste diferențiate (9) și capilare sanguine (10); 11 - osteoblaste; 12 - țesut osos format endocondral; 13 - punct de osificare în epifiză
Dezvoltarea osului în locul cartilajului, adică osteogeneza indirectă, începe în zona diafizei (osificarea pericondală). Educaţie inel osos pericondral(manșetă) este precedată de proliferarea vaselor de sânge cu diferențiere în pericondrul adiacent părții medii a diafizei osteoblastelor, care formează mai întâi o manșetă. țesut osos fibros grosier(centrul primar de osificare), apoi înlocuit lamelar.
Formarea unei manșete osoase perturbă nutriția cartilajului. Ca urmare, apar modificări distrofice în centrul părții diafizare a rudimentului cartilaginos. Condrocitele devin vacuolizate, nucleele lor devin picnotizate și se formează așa-numitele condrocite veziculare. Creșterea cartilajului în acest loc se oprește. Expansiunea inelului osos pericondral este însoțită de o creștere a zonei de distrugere a cartilajului și de apariția osteoclastelor, care eliberează căile pentru vasele de sânge și osteoblastele care cresc în modelul osos tubular (vezi Fig. 8.30). Aceasta duce la apariția focarelor de osificare endocondrală (centri de osificare secundare). Datorită creșterii continue a secțiunilor distale neschimbate adiacente ale diafizei, condrocitele de la marginea epifizei și diafizei sunt colectate în coloane, a căror direcție coincide cu axa lungă a viitorului os. Astfel, în coloana de condrocite există două procese direcționate opus - reproducerea și creșterea în părțile distale ale diafizei și procesele distrofice în partea sa proximală. În același timp, între celulele umflate se depun săruri minerale, determinând apariția unei bazofilii severe și a fragilității cartilajului.
Din momentul în care rețeaua vasculară crește și apar osteoblastele, pericondrul este reconstruit, transformându-se în periost. Ulterior, vasele de sânge cu mezenchim înconjurător, celule osteogene și osteoclaste cresc prin deschiderile manșetei osoase și intră în contact cu cartilajul calcificat (vezi Fig. 8.30). Sub influența enzimelor secretate de osteoclaste are loc dizolvarea (condroliza) substanței intercelulare calcificate. Cartilajul diafizar este distrus, în el apar spații alungite, în care osteoblastele „se așează”, formând țesut osos pe suprafața zonelor rămase ale substanței intercelulare calcificate a cartilajului.
Centru de osificare primar sau diafizar. Procesul de formare a osului în interiorul rudimentului cartilaginos se numește osificare endocondrală (greacă. endon- interior).
Concomitent cu dezvoltarea osului endocondral, apar semne ale distrugerii acestuia de către osteoclaste. Datorită distrugerii țesutului osos endocondral, se formează cavități și spații și mai mari (cavități de resorbție) și, în final, o cavitatea măduvei osoase. Din mezenchimul care pătrunde aici se formează stroma măduvei osoase, în care se instalează celulele stem din sânge și țesut conjunctiv. În același timp, din ce în ce mai multe noi bare transversale de țesut osos apar de-a lungul periferiei diafizei din partea periostului. Țesutul osos, crescând în lungime spre epifize și crescând în grosime, formează un strat dens de os.
Organizarea osului periostal decurge diferit de organizarea țesutului osos endocondral. În locul osului fibros grosier care se prăbușește, în jurul vaselor care au pătruns aici, care se desfășoară de-a lungul axei lungi a rudimentului osos, încep să se formeze plăci concentrice, formate din fibre subțiri de colagen orientate paralel și o substanță intercelulară cimentantă. Acesta este cum osteone primare. Lumenul lor este larg, limitele plăcilor nu sunt conturate ascuțit. În urma apariției primei generații de osteoni, dezvoltarea de plăci de brâu extern (general),înconjoară osul în zona diafizei. În urma diafizei, în epifize apar centrii de osificare. Aceasta este mai întâi precedată de diferențierea condrocitelor, hipertrofia lor, urmată de deteriorarea nutriției, degenerarea și calcificarea substanței intercelulare. În viitor, se remarcă un proces de osificare similar celui descris mai sus. Osificarea este însoțită de creșterea în interiorul epifizelor vaselor.
În zona intermediară dintre diafize și epifize se păstrează țesutul cartilajului - cartilaj metafizar, care este o zonă de creștere a osului în lungime.
Structura histologică a osului tubular ca organ
Osul tubular ca organ este construit în principal din țesut osos lamelar, cu excepția tuberculilor. Exteriorul osului este acoperit de periost, cu excepția suprafețelor articulare ale epifizelor, care sunt acoperite cu un tip de cartilaj hialin.

Orez. 8.31. Structura osului tubular (conform lui V. G. Eliseev, Yu. I. Afanasiev, E. F. Kotovsky):
A- periostul; b- substanță osoasă compactă; V- endostul; G- cavitatea măduvei osoase. 1 - un strat de plăci comune externe; 2 - osteon; 3 - canal osteon; 4 - plăci de inserare; 5 - un strat de plăci interne comune; 6 - tra-becula osoasa a substantei spongioase; 7 - stratul fibros al periostului; 8 - vasele de sânge ale periostului; 9 - canal perforant; 10 - osteocite
Periost, sau periost. Periostul are două straturi: exterior (fibros) și interior (celular). Stratul exterior este format în principal din țesut conjunctiv fibros (Fig. 8.31, 8.32). Stratul interior conține un număr mare de celule: celule cambiale, preosteoblaste și osteoblaste cu diferite grade de diferențiere. Celulele cambiale în formă de fus au un volum mic de citoplasmă și un aparat sintetic dezvoltat moderat. Preosteoblastele sunt celule de formă ovală cu proliferare viguroasă, capabile să sintetizeze glicozaminoglicani. Osteoblastele sunt caracterizate de un aparat de sinteză a proteinelor (colagen) bine dezvoltat. Vasele și nervii care alimentează osul trec prin periost.
Periostul conectează osul cu țesuturile din jur și participă la trofismul, dezvoltarea, creșterea și regenerarea acestuia.
Structura diafizei. Substanța compactă care formează diafiza osoasă este formată din plăci osoase, a căror grosime variază de la 4 la 12-15 microni. Plăcile osoase sunt aranjate într-o anumită ordine,

Orez. 8.32. Periostul (după Yu. I. Afanasiev):
1 - strat exterior (fibros); 2 - stratul interior (celular); 3 - celule osteogene; 4 - țesut osos
formând sisteme complexe. Există trei straturi în diafiză: exterior un strat de plăci de brâu (general, general), in medie, format din plăci osoase stratificate concentric în jurul vaselor - osteoni și numit strat de osteon (Fig. 8.33) și interior strat de plăci de brâu (comune).
Plăcile exterioare nu formează inele complete în jurul diafizei osoase; ele sunt suprapuse la suprafață de straturi ulterioare de plăci. Plăcile interne sunt bine dezvoltate numai acolo unde substanța osoasă compactă mărginește direct cavitatea medulară. În aceleași locuri în care substanța compactă trece în substanța spongioasă, plăcile sale comune interne continuă în plăcile trabeculelor substanței spongioase.
Plăcile exterioare sunt străpunse canale perforante (Volkmann), prin care intră vase din periost în os. Din periost, fibrele de colagen pătrund în os sub diferite unghiuri. Aceste fibre sunt numite fibre perforante (Sharpey). Cel mai adesea se ramifică numai în stratul exterior al laminelor comune, dar pot pătrunde și în stratul osteonic mijlociu, dar nu intră niciodată în laminele osteonice.
În stratul mijlociu sunt oase plăci concentrice (osteonice). formează osteoni. Între osteoni sunt situate plăci intermediare (inserate). Grosimea și lungimea plăcilor osoase variază de la câteva zeci la sute de micrometri. Osteoni(Sistemele Haversiene) sunt unități structurale ale substanței compacte a osului tubular (vezi Fig. 8.31, Fig. 8.33). Sunt formațiuni cilindrice formate din plăci osoase concentrice, parcă introduse unele în altele. În plăcile osoase și între ele se află corpurile celulelor osoase și procesele lor, înfundate în substanța intercelulară osoasă. Fiecare osteon este delimitat de osteonii vecini printr-o linie de cimentare (fuziune) formată din substanța fundamentală. In central

Orez. 8.33. Osteon:
A- microfotografie (colorare prin metoda Schmorl); b- microscopia electronică cu baleiaj a unui fragment de os (pregătire de O. V. Slesarev). 1 - canal osteon; 2 - osteocite (lacune - b); 3 - plăci osoase
Canalul osteon conține vase de sânge cu țesut conjunctiv însoțitor și celule osteogene.
În diafiza unui os lung, osteonii sunt localizați predominant paralel cu axul lung. Canalele osteonice se anastomozează între ele, la locurile anastomozei plăcile adiacente își schimbă direcția (vezi Fig. 8.31). Astfel de canale sunt numite perforare, sau hrănitoare. Vasele situate in canalele osteonului comunica intre ele si cu vasele maduvei osoase si periostului. Cea mai mare parte a diafizei este alcătuită din substanța compactă a oaselor tubulare. Pe suprafața interioară a diametrului
physis mărginind cavitatea medulară, țesutul osos lamelar formează barele transversale osoase ale osului spongios. Cavitatea diafizei oaselor tubulare este umplută cu măduvă osoasă.
Endostul - membrana care acoperă osul din partea laterală a cavității medulare. În endostul suprafeței osoase formate se distinge o linie osmiofilă pe marginea exterioară a substanței osoase mineralizate; stratul osteoid, format dintr-o substanță amorfă, fibrile de colagen și osteoblaste, capilare sanguine și terminații nervoase, un strat de celule care separă vag endostul de elementele măduvei osoase. Grosimea endostului depășește 1-2 microni, dar este mai mică decât cea a periostului.
În zonele de formare osoasă activă, grosimea endostului crește de 10-20 de ori datorită stratului osteoid datorită activității sintetice crescute a osteoblastelor și a precursorilor acestora. În timpul remodelării osoase, în endost se găsesc osteoclaste. În endostul osului îmbătrânit, populația de osteoblaste și celule progenitoare scade, dar activitatea osteoclastelor crește, ceea ce duce la subțierea stratului compact și la restructurarea osului spongios.
Intre endost si periost exista o anumita microcirculatie a lichidului si a mineralelor datorita sistemului lacunar-tubular al tesutului osos.
Vascularizarea țesutului osos. Vasele de sânge formează o rețea densă în stratul interior al periostului. De aici provin ramuri arteriale subțiri, care furnizează osteonii cu sânge prin deschideri de nutrienți, apoi pătrund în măduva osoasă și participă la formarea rețelei capilare care o hrănește. Vasele limfatice sunt localizate în principal în stratul exterior al periostului.
Inervația țesutului osos.În periost, fibrele nervoase mielinizate și nemielinice formează un plex. Unele dintre fibre însoțesc vasele de sânge și pătrund odată cu ele prin deschiderile de nutrienți în canalele cu același nume, apoi în canalele osteonului și apoi ajung în măduva osoasă. O altă parte a fibrelor se termină în periost cu ramuri nervoase libere și, de asemenea, participă la formarea corpurilor încapsulate.
Creșterea oaselor tubulare. Creșterea osoasă este un proces foarte lung. Începe la om încă din fazele embrionare timpurii și se termină în medie la vârsta de 20 de ani. Pe toată perioada de creștere, osul crește atât în lungime, cât și în lățime. Creșterea în lungime a osului tubular este asigurată de prezența unei plăci de creștere cartilaginoasă metaepifizară, în care se manifestă două procese histogenetice opuse.
Una este distrugerea plăcii epifizare, iar cealaltă, opusă, este refacerea constantă a țesutului cartilajului prin formarea de noi celule. Cu toate acestea, în timp, procesele de distrugere a celulelor încep să prevaleze asupra proceselor de neoplasm, în urma cărora placa cartilaginoasă devine mai subțire și dispare. Creșterea osoasă în lungime se oprește.
În cartilajul metaepifizar există frontieră zonă, zonă celule columnare si zona celule veziculare. Zona de frontieră situată în apropiere
epifiza, constă din celule rotunde și ovale și grupuri izogenice simple care asigură legătura între placa cartilaginoasă și osul epifizei. În cavitățile dintre os și cartilaj există capilare sanguine care furnizează nutriție celulelor din zonele mai profunde ale plăcii cartilaginoase. Zona celulară columnară conține celule care proliferează activ care formează coloane de-a lungul axei longitudinale a osului și asigură creșterea și lungimea acestuia. Capetele proximale ale coloanelor constau din celule cartilaginoase maturizate, diferențiate. Sunt bogate în glicogen și fosfatază alcalină. Ambele zone sunt cele mai reactive la acțiunea hormonilor și a altor factori care influențează procesele de osificare și creșterea osoasă. Zona celulelor veziculare se caracterizează prin hidratarea și distrugerea condrocitelor urmată de osificare endocondrală. Partea distală a acestei zone mărginește diafiza, de unde capilarele sanguine și celulele osteogene pătrund în ea. Coloanele orientate longitudinal ale osului endocondral sunt în esență tuburi osoase în care se formează osteoni.
Ulterior, centrii de osificare din diafiză și epifiză se contopesc, iar creșterea osoasă în lungime se termină.
Creșterea osului tubular în lățime se datorează periostului. Pe partea laterală a periostului încep să se formeze foarte devreme straturi concentrice de țesut osos lamelar. Acest apozițional creșterea continuă până când formarea osului este completă. Numărul de osteoni imediat după naștere este mic, dar până la vârsta de 25 de ani numărul lor în oasele lungi ale membrelor crește semnificativ.
Regenerare. Regenerarea fiziologică a țesutului osos are loc lent datorită celulelor osteogene ale periostului - endost și celulelor osteogene din canalul osteon. Regenerarea post-traumatică a țesutului osos decurge mai bine în cazurile în care capetele osului rupt nu sunt deplasate unul față de celălalt. Procesul de osteogeneză este precedat de formarea calusului de țesut conjunctiv, în grosimea căruia se pot forma insule cartilaginoase (Fig. 8.34). Osificarea în acest caz are loc în funcție de tipul de osteogeneză secundară (indirectă). În condiții de oxigenare optimă a țesuturilor, repoziționare și fixare bună a capetelor osului rupt, regenerarea are loc fără formarea calusului. Cu toate acestea, înainte ca osteoblastele să înceapă să construiască os, osteoclastele formează un mic spațiu între capetele de împerechere ale osului. Acest model biologic stă la baza utilizării de către traumatologi a dispozitivelor pentru întinderea treptată a oaselor topite pe întreaga perioadă de regenerare.
Remodelarea osoasă și factorii care influențează structura acestuia
În țesutul osos de-a lungul vieții unei persoane, au loc procese interconectate de distrugere și creație, cauzate de sarcini funcționale și de alți factori ai mediului extern și intern. Restructurarea osteonilor este întotdeauna asociată cu distrugerea osteonilor primari și formarea simultană de noi osteoni atât la locul distrugerii, cât și pe partea laterală a periostului. Sub influența osteoclastelor activate

Orez. 8.34. Regenerarea posttraumatică a osului tubular: A- localizarea leziunii; dumnezeu- etape succesive de regenerare fără fixare rigidă a oaselor reduse (b 1, în 1- fragmente); d- regenerare după fixarea fragmentelor. 1 - periost; 2 - bare transversale din țesut osos din fibre grosiere; 3 - tesutul conjunctiv se regenereaza cu insule de tesut cartilaginos; 4 - os regenerat din țesut osos fibros grosier; 5 - linie de fuziune (după R.V. Krstic, cu modificări)
diverși factori, plăcile osoase ale osteonului sunt distruse, iar în locul ei se formează o cavitate. Acest proces se numește resorbţie(din lat. resorbție- resorbţia) ţesutului osos. Osteoblastele apar în cavitatea rezultată din jurul vasului rămas și începe construcția de noi plăci, stratificate concentric una peste alta. Așa apar generațiile secundare de osteoni. Între osteoni se află rămășițele de osteoni distruși din generațiile anterioare. Procesul de restructurare a osteonului nu se oprește nici după sfârșitul creșterii osoase.
Printre factorii care influențează restructurarea țesutului osos, așa-numitul efect piezoelectric joacă un rol semnificativ. S-a dovedit că la îndoirea plăcii osoase, apare o anumită diferență de potențial între părțile concave și convexe. Prima este încărcată negativ,
iar al doilea este pozitiv. Pe o suprafață încărcată negativ se observă întotdeauna activarea osteoblastelor și procesul de formare nouă apozițională a țesutului osos, în timp ce pe o suprafață încărcată pozitiv, dimpotrivă, se observă resorbția acestuia cu ajutorul osteoclastelor. Crearea artificială a unei diferențe de potențial duce la același rezultat (Fig. 8.35). Potențialul zero, lipsa stresului fizic asupra țesutului osos (imobilizare prelungită, aflarea în stare de imponderabilitate etc.) determină creșterea funcțiilor osteoclastelor și excreția de săruri.
Structura țesutului osos și a oaselor este influențată de vitamine (C, A, D), hormoni ai tiroidei, paratiroidei și a altor glande endocrine.
În special, cu o cantitate insuficientă de vitamina C în organism (de exemplu, cu scorbut), formarea fibrelor de colagen este suprimată, activitatea osteoblastelor este slăbită, iar activitatea lor fosfatază este redusă, ceea ce duce practic la încetarea creșterea osoasă datorită inhibării formării bazei organice a țesutului osos. Cu deficit de vitamina D (rahitism), nu are loc calcificarea completă a matricei osoase organice, ceea ce determină înmuierea oaselor (osteomalacie). Vitamina A susține creșterea oaselor, dar un exces al acestei vitamine crește distrugerea cartilajului metaepifizar de către osteoclaste - zona de creștere a oaselor - și încetinește alungirea acestora.
Cu un exces de hormon paratiroidian - paratirina - se observă o creștere a activității osteoclastelor și a resorbției osoase. Tirocalcitonina, produsă de celulele C ale glandei tiroide, acționează diametral opus, reducând funcția osteoclastelor care au receptori pentru acest hormon. Cu hipofuncția glandei tiroide, creșterea oaselor tubulare lungi încetinește ca urmare a suprimării activității osteoblastelor și a inhibării procesului de osificare. Regenerarea osoasă în acest caz are loc slab și incomplet. În cazul subdezvoltării testiculare sau al castrării prepuberale, osificarea plăcii metaepifizare este întârziată, drept urmare brațele și picioarele unui astfel de individ devin disproporționat de lungi. Cu o lipsă de estrogen după menopauză, femeile dezvoltă uneori osteoporoză. Odată cu pubertatea timpurie, se așteaptă încetarea creșterii din cauza fuziunii premature diafizar-epifizare a oaselor. Un anumit rol pozitiv în creșterea osoasă îl joacă hormonul somatotrop al adenohipofizei, care stimulează dezvoltarea proporțională a scheletului la vârste tinere (adolescente) și disproporționat (acromegalia) la adulți.
Modificări de vârstă.Țesuturile conjunctive suferă modificări ale structurii, cantității și compoziției chimice odată cu vârsta. Odată cu vârsta, masa totală a formațiunilor de țesut conjunctiv și creșterea scheletului osos cresc. În multe tipuri de structuri ale țesutului conjunctiv, raportul dintre tipurile de colagen și glicozaminoglicani se modifică; în special, conțin mai mulți compuși sulfatați.
Conexiuni osoase
Două oase pot avea conexiuni continuu(sindesmoze, sincondroze și sinostoze) și intermitent(articulații).
Conexiuni continue- legături cu ajutorul țesutului conjunctiv fibros dens, din care mănunchiuri sub formă de fibre perforante sunt introduse în os;

Orez. 8.35. Efect piezoelectric (explicații în text): I - diagrama organizării structurale a trabeculei osoase; II - activarea osteoclastelor și osteoblastelor la modificarea formei trabeculei osoase; III - crearea artificială a diferenței de potențial (conform lui Yu. I. Afanasyev)
tesatura noua. Un exemplu de astfel de conexiuni sunt suturile oaselor parietale ale craniului, membrana de țesut conjunctiv dintre oasele radius și ulna.
Sincondroze (simfize)- conexiuni realizate de cartilaj, precum discurile intervertebrale. Ele constau dintr-un inel fibros exterior și o parte interioară numită nucleu pulpos. Ambele părți sunt vag separate și se transformă imperceptibil una în cealaltă. Nucleul pulpos este situat în zona interioară a discului intervertebral. La diferite perioade de vârstă are structură diferită. La vârsta de până la 2 ani, este o cavitate cu conținut omogen, care conține doar celule individuale. În anii următori de viață, această cavitate este împărțită în camere separate. Începând cu vârsta de 6-8 ani, în nucleul pulpos se constată apariția și apoi creșterea numărului de fibre de colagen și celule de cartilaj. De la vârsta de 15 ani, creșterea fibrelor și a celulelor cartilajului se intensifică și mai mult, iar la vârsta de 20-23 de ani, nucleul pulpos capătă aspectul caracteristic de cartilaj fibros. Un exemplu de altă conexiune, mai strânsă, ar fi simfiza pubiană. Sincondrozele includ și conexiuni ale epifizei și diafizei cu ajutorul cartilajului metaepifizar.
Sinostoza- articulații strânse ale oaselor fără țesut conjunctiv fibros, cum ar fi oasele pelvine.
Articulațiile intermitente sau articulațiile (diartroză) constau din suprafețe articulare acoperite cu cartilaj și, în unele cazuri, un menisc intermediar cartilaginos și o capsulă articulară. Capsula articulară este formată dintr-un strat fibros exterior și un strat sinovial interior. Acesta din urmă este înțeles ca un strat de țesut conjunctiv diferențiat specific, care conține vase sanguine și limfatice, fibre nervoase și terminații. Poziția limită a acestui țesut conjunctiv, neobișnuită pentru alți derivați mezenchimatoși, întinderea constantă, deplasarea și presiunea datorită participării funcției locomotorii a articulației determină creșterea și caracteristicile sale structurale.
În membrana sinovială a mamiferelor și a omului se disting două straturi fibroase colagen-elastice (superficial și profund) și un strat de acoperire care căptușește cavitatea (vezi Fig. 8.20). Nu există o limită ascuțită între straturi. In articulatiile mari exista un strat subsinovial bogat in tesut adipos, marginind capsula fibroasa. Colagenul și fibrele elastice ale stratului de suprafață sunt orientate în direcția axei lungi a articulației. În stratul profund sunt situate în unghi față de fibrele stratului de suprafață.
Stratul de acoperire al membranei sinoviale este format din celule - sinoviocite. Există sinoviocite macrofage și fibroblaste sinoviale, care au capacitatea de a produce și secreta acid hialuronic, o componentă specifică a lichidului sinovial.
Vasele de sânge pătrund în sinoviale din țesuturile subiacente și sunt distribuite pe toată grosimea acestuia, inclusiv în stratul tegumentar, unde sunt situate direct sub sinoviocite. Astfel, cavitatea sinovială este separată de fluxul sanguin numai de celule, principala substanță a țesutului conjunctiv și de endoteliul capilarelor în sine. Endoteliul hemocapilarelor membranelor sinoviale se caracterizează prin fenestre și capacitatea de fagocitare. Capilarele limfatice sunt întotdeauna situate mai adânc decât capilarele sanguine în stratul fibros superficial.
Membrana sinovială este bogat inervată de fibre de natură aferentă și eferentă (simpatică).
Întrebări de control
1. Mezenchimul ca sursă de dezvoltare a țesuturilor conjunctive: conceptul de diferențiere divergentă a mezenchimului, caracteristicile morfologice ale mezenchimului.
2. Tesuturi conjunctive cu proprietati deosebite: clasificare, topografie in organism, structura, functii.
3. Diferențele celulare ale țesutului conjunctiv lax: surse de dezvoltare, structură, funcții, participare la regenerarea fiziologică și reparatorie.
4. Tesutul cartilaginos: clasificare, topografie, structura, functii, regenerare.
5. Țesutul osos: osteogeneză directă și indirectă, structură, compoziție celular-diferențială, regenerare.
Histologie, embriologie, citologie: manual / Yu. I. Afanasyev, N. A. Yurina, E. F. Kotovsky etc. - Ed. a VI-a, revizuită. si suplimentare - 2012. - 800 p. : bolnav.