結合組織の構造の特徴。 結合組織 (人体解剖学)。 結合組織の構造
結合組織胚の発生中に間葉から形成されます。 体内で多くの重要な機能を果たします。 - サポートメカニカル- 結合組織が体全体(骨格)およびほとんどの臓器の支持フレームであるという事実による。
- 栄養(代謝)-結合組織はどこにでもある血管(たとえ最も小さな血管であっても)に付随しており、血液と臓器の他の組織の間の代謝プロセスの実行における仲介者であるという事実によって決定されます。
- 保護- 免疫反応に積極的に関与する結合組織内の食細胞の存在に依存します。
- 修復物(プラスチック)- 再生プロセス(損傷または疾患後の組織および器官の完全性の回復)における結合組織の積極的な関与によって現れます。
結合組織は体重の半分以上を占め、脊椎動物の進化の過程でその発達の程度は徐々に増加します。 学者によると A.A.ボゴモレッツ。 結合組織は単一のシステムを形成しており、そのシステムは広範囲に分布しており、細胞の急速な複製と体の目的の部分への移動、それらの相互作用、再生現象や免疫反応への積極的な参加を特徴としています。
結合組織の構造の特徴は、細胞とともに、非常に発達した細胞間物質(非晶質物質と繊維)がその中に存在することです。 細胞間物質の構造に基づいて、結合組織の主な種類を区別できます。
血液とリンパ
血液(血漿)やリンパの細胞間物質には繊維構造が存在しないため、これらの種類の結合組織は液体の粘稠度を持っています。 リンパの化学組成は血漿に近く、赤血球(赤血球)、白血球(白血球)、血小板(血小板)といったいわゆる有形成分が含まれています。 哺乳類では、列挙された形状要素のうち、同じであるのは白血球だけです。 赤血球は、核を有する細胞から発生し、ヘモグロビンの蓄積の過程で核を失うため、後細胞形成で表されます。 哺乳動物の血小板は、主に赤色骨髄に存在する特殊な巨細胞 (巨核球) の断片です。 白血球は顆粒球(顆粒球)と非顆粒球(無顆粒球)に分けられます。 次に、顆粒球は酸性塩基または中性色素に関連しているため、好酸性、好塩基性、好中性の 3 つに分類されます。 無顆粒球はリンパ球と単球に分化します。血液の主な機能は輸送であり、その発現は栄養、呼吸、保護、恒常性です。
網様結合組織
網様結合組織(ラテン語のReticulumに由来)は、バケツの形をしており、突起で互いに接触している細胞で構成されているため、メッシュ構造を持っています。その細胞の中には、細胞間物質を生成する線維芽細胞と、血液単球から形成される固定マクロファージが含まれます。 網状組織の細胞間物質は、コラーゲンの一種である非晶質物質と細い網状線維で代表されます。 網状組織は造血器官の一部であり、半液体の粘度を持っています。 造血に関与し、血球が発達するための環境(微環境)を作成し、食細胞の助けを借りて保護機能を果たします。
線維性結合組織
線維性結合組織では、線維の数は中程度(疎な線維組織)である場合もあれば、より多く(密な線維組織)である場合もあります。 疎性線維性結合組織は、最も一般的な種類の結合組織です。 それは主に栄養機能と保護機能を実行し、体内で起こるほぼすべての生理学的反応および保護反応に関与します。 疎性結合組織の細胞には、線維芽細胞、大食細胞、形質細胞 (形質細胞)、組織好塩基球、脂肪細胞 (脂肪細胞)、および色素細胞 (色素細胞) が含まれます。結合組織細胞の最も多くのグループは線維芽細胞であり、その中間物質 (非晶質物質と線維) を形成します。 線維芽細胞は、分泌細胞に典型的な特殊な超微細構造を持っています(よく発達した顆粒小胞体とゴルジ複合体を含んでいます)。 若い線維芽細胞は分裂することができ、バケツの形をしています。 成熟した線維芽細胞(線維細胞)は分裂できません。 細胞小器官の大部分が減少し、その機能活性が大幅に低下します。
結合組織細胞の 2 番目に大きなグループは大食細胞です。 それらは血液単球から形成され、組織マクロファージへの変換の過程で細胞小器官およびリソソームの含有量が増加します。 これらの変化は、組織マクロファージの活発な食作用と多くの生物学的に活性な物質の合成能力を反映しています。
形質細胞 Bリンパ球から形成されます。 それらは、円形または楕円形、放射状に配向したヘテロクロマチンパッチを備えた偏心して配置された核、よく発達した顆粒小胞体および核の近く(顆粒小胞体を失った細胞質の領域)に位置するゴルジ複合体を備えています。 。 これらの細胞の機能は、体内の抗原を中和する特別なタンパク質(ガンマグロブリン)である抗体の合成です。
組織の好塩基球は細胞質に好塩基性顆粒を含んでいます。 これらの細胞は、いわゆる生体アミン (ヘパリン、ヒスタミン、セロトニン) を分泌し、血液凝固、小血管の組織透過性などの調節に関与します。この点で、組織好塩基球は局所恒常性の調節因子です。
脂肪細胞(脂肪細胞)は、エネルギーおよび栄養物質、断熱物質である脂肪を蓄積することができます。 白色脂肪と褐色脂肪の脂肪細胞があります。 白色脂肪脂肪細胞には、中性脂肪によって形成された 1 つの大きな脂肪封入体 (液滴) があり、核を含む細胞質の大部分が細胞の極の 1 つに押し込まれています。
褐色脂肪脂肪細胞はサイズが小さいです。 それらの核は中心に位置し、脂肪は多数の小さな滴の形で細胞質にあります。 褐色脂肪細胞はミトコンドリアを多く含んでいるのが特徴です。 褐色脂肪組織の主な機能は熱を生成することです。 人間の場合、生後数か月間で十分に発達し、その後白色脂肪組織に置き換わります。
色素細胞(色素細胞) - 色素顆粒を含むバケツ状の細胞要素 - 細胞質内のメラニン。 人間および哺乳動物では、それらは皮膚(真皮と表皮の両方)、毛包、髄膜、虹彩および脈絡膜の一部です。 これらの細胞は保護機能を果たし、皮膚や視覚器官に対する太陽光の有害な影響を軽減します。
組織は、同じ構造、機能、起源を持つ細胞と細胞間物質の集合です。
哺乳類と人間の体内では、4 種類の組織が区別されます。上皮組織、結合組織では、骨、軟骨、脂肪組織が区別されます。 筋肉質で神経質。
組織 - 体内の位置、種類、機能、構造
組織は、同じ構造、起源、機能を持つ細胞と細胞間物質のシステムです。
細胞間物質は細胞活動の産物です。 細胞間のコミュニケーションを提供し、細胞にとって好ましい環境を作り出します。 血漿などの液体の場合もあります。 非晶質 - 軟骨; 構造化された - 筋繊維。 硬骨組織(塩の形)。
組織細胞にはさまざまな形状があり、それによって機能が決まります。 生地は以下の4種類に分けられます。
- 上皮 - 境界組織: 皮膚、粘膜。
- 結合 - 私たちの体の内部環境。
- 筋;
- 神経組織。
上皮組織
上皮(境界)組織 - 体の表面、すべての内臓と体の空洞の粘膜、漿液性膜を覆い、外分泌腺と内分泌腺も形成します。 粘膜の内側を覆う上皮は基底膜上に位置し、その内面は直接外部環境に面しています。 その栄養は、基底膜を通した血管からの物質と酸素の拡散によって達成されます。
特徴:細胞数が多く、細胞間物質が少なく、基底膜に代表される。
上皮組織は次の機能を実行します。
- 保護;
- 排泄物;
- 吸引
上皮の分類。 層の数に基づいて、単層と多層が区別されます。 形状に応じて、平面、立方体、円筒形に分類されます。
すべての上皮細胞が基底膜に到達している場合、それは単層上皮であり、1 つの列の細胞のみが基底膜に接続され、他の細胞が自由である場合、それは多層上皮です。 単層上皮は、核の位置のレベルに応じて、単列または複数列になることがあります。 場合によっては、単核または多核上皮が外部環境に面した繊毛を持つこともあります。
重層上皮 上皮(外皮)組織、または上皮は、体の外皮、すべての内臓および空洞の粘膜を裏打ちする細胞の境界層であり、また多くの腺の基礎も形成します。
腺上皮 上皮は生体 (内部環境) を外部環境から分離しますが、同時に生体と環境の相互作用の仲介者としても機能します。 上皮細胞は互いに緊密に結合しており、微生物や異物の体内への侵入を防ぐ機械的障壁を形成しています。 上皮組織細胞の生存期間は短く、すぐに新しいものに置き換わります (このプロセスは再生と呼ばれます)。
上皮組織は、分泌 (外分泌腺および内分泌腺)、吸収 (腸上皮)、ガス交換 (肺上皮) など、他の多くの機能にも関与しています。
上皮の主な特徴は、それが高密度に詰まった細胞の連続層で構成されていることです。 上皮は、体のすべての表面を覆う細胞層の形、および細胞の大きな蓄積の形、つまり肝臓、膵臓、甲状腺、唾液腺などの腺の形をとることができます。最初のケースでは、上皮は上にあります。基底膜は、上皮をその下にある結合組織から分離します。 ただし、例外もあります。リンパ組織の上皮細胞は結合組織の要素と交互に存在し、そのような上皮は非定型と呼ばれます。
層状に配置された上皮細胞は、多くの層に存在する場合 (重層上皮)、または 1 つの層に存在する場合 (単層上皮) があります。 細胞の高さに基づいて、上皮は平板状、立方体状、角柱状、円筒状に分類されます。
単層扁平上皮 - 漿膜の表面を覆っています:胸膜、肺、腹膜、心臓の心膜。
単層立方上皮 - 尿細管の壁と腺の排泄管を形成します。
単層円柱上皮 - 胃粘膜を形成します。
境界上皮(細胞の外表面には栄養素の吸収を確実にする微絨毛によって形成された境界がある単層の円筒上皮)は、小腸の粘膜の内側を覆っています。
繊毛上皮(繊毛上皮)は、円筒形の細胞からなる偽重層上皮であり、その内端、つまり腔または管に面した部分には、常に振動する毛状構造(繊毛)が装備されています。繊毛は卵子の動きを保証します。チューブ。 気道から細菌や塵を除去します。
重層上皮は身体と外部環境の境界に位置します。 上皮で角化プロセスが発生する場合、つまり細胞の上層が角質スケールに変化する場合、そのような多層上皮は角化(皮膚表面)と呼ばれます。 多層上皮は、口、食道、目の角膜の粘膜を覆っています。
移行上皮は膀胱、腎盂、尿管の壁を覆っています。 これらの器官が満たされると、移行上皮が伸び、細胞がある列から別の列に移動できるようになります。
腺上皮 - 腺を形成し、分泌機能を実行します(放出物質 - 外部環境に排泄されるか、血液やリンパ(ホルモン)に入る秘密)。 体の生命活動に必要な物質を生成および分泌する細胞の能力は、分泌と呼ばれます。 この点において、そのような上皮は分泌上皮とも呼ばれた。
結合組織
結合組織 細胞、細胞間物質、結合組織線維から構成されます。 それは骨、軟骨、腱、靱帯、血液、脂肪で構成され、いわゆる臓器の間質(骨格)の形ですべての臓器(疎性結合組織)に存在します。
上皮組織とは対照的に、すべての種類の結合組織(脂肪組織を除く)では、細胞間物質の体積が細胞よりも多く、つまり細胞間物質が非常によく発現されています。 細胞間物質の化学組成と物理的特性は、結合組織の種類によって非常に多様です。 たとえば、血液 - 細胞間物質がよく発達しているため、その中の細胞は「浮遊」し、自由に動きます。
一般に、結合組織はいわゆる体の内部環境を構成します。 それは非常に多様であり、密集したものから緩いもの、液体中に細胞が存在する血液やリンパ液まで、さまざまな種類で表されます。 結合組織の種類の基本的な違いは、細胞成分の比率と細胞間物質の性質によって決まります。
高密度の線維性結合組織 (筋腱、関節靱帯) は線維構造が大半を占めており、重大な機械的ストレスを受けます。
緩い線維性結合組織は体内で非常に一般的です。 それどころか、さまざまな種類の細胞形態が非常に豊富です。 それらのうちのいくつかは組織線維 (線維芽細胞) の形成に関与しており、その他は特に重要であり、免疫機構 (マクロファージ、リンパ球、組織好塩基球、形質細胞) を介したものなど、主に保護および制御プロセスを提供します。
骨
骨組織 骨格の骨を形成する骨組織は非常に耐久性があります。 体型(体質)を維持し、頭蓋骨、胸腔、骨盤腔にある臓器を保護し、ミネラル代謝に関与します。 組織は細胞(骨細胞)と細胞間物質で構成されており、その中に血管を伴う栄養チャネルが存在します。 細胞間物質には最大 70% の無機塩 (カルシウム、リン、マグネシウム) が含まれています。
骨組織はその発達において、線維性段階と層状段階を経ます。 骨のさまざまな部分で、骨は緻密なまたは海綿状の骨物質の形で組織されます。
軟骨組織
軟骨組織は細胞(軟骨細胞)と細胞間物質(軟骨基質)で構成されており、弾力性が高いのが特徴です。 軟骨の大部分を形成するため、サポート機能を果たします。
軟骨組織には 3 種類あります。硝子は、気管、気管支、肋骨の端、骨の関節面の軟骨の一部です。 弾性があり、耳介と喉頭蓋を形成します。 繊維状で、椎間板と恥骨の関節にあります。
脂肪組織
脂肪組織は疎性結合組織に似ています。 細胞は大きく、脂肪で満たされています。 脂肪組織は、栄養、形状形成、体温調節の機能を果たします。 脂肪組織は白色と褐色の2種類に分けられます。 ヒトでは、白色脂肪組織が優勢で、その一部は臓器を取り囲んで、人体内での位置やその他の機能を維持しています。 人間の褐色脂肪組織の量は少ないです(主に新生児に見られます)。 褐色脂肪組織の主な機能は熱生成です。 褐色脂肪組織は、冬眠中の動物の体温と新生児の体温を維持します。
筋
筋細胞は常に一方向に伸びているため、筋線維と呼ばれます。
筋肉組織の分類は、組織の構造(組織学的)に基づいて行われます:横縞の有無、および収縮のメカニズム - 随意的(骨格筋など)または不随意的(平滑筋)または心筋)。
筋肉組織は興奮性を持ち、神経系や特定の物質の影響下で積極的に収縮する能力があります。 顕微鏡的な違いにより、この組織の2つのタイプ、つまり平滑(横紋のない)と横紋のある(横紋のある)を区別することができます。
平滑筋組織は細胞構造を持っています。 内臓(腸、子宮、膀胱など)、血管、リンパ管の壁の筋肉膜を形成します。 その収縮は無意識に起こります。
横紋筋組織は筋線維で構成されており、それぞれの筋線維は数千の細胞で構成され、核に加えて融合して 1 つの構造になっています。 骨格筋を形成します。 自由に短くすることができます。
さまざまな横紋筋組織が心筋であり、独特の能力を持っています。 一生(約70年)の間に、心筋は250万回以上収縮します。 これほどの強度を備えた生地は他にありません。 心筋組織には横縞があります。 ただし、骨格筋とは異なり、筋線維が交わる特別な領域があります。 この構造により、1本の繊維の収縮がすぐに隣の繊維に伝わります。 これにより、心筋の大部分が同時に収縮します。
また、筋肉組織の構造上の特徴は、その細胞にアクチンとミオシンという 2 つのタンパク質によって形成された筋原線維の束が含まれていることです。
神経組織
神経組織は、神経 (ニューロン) とグリアという 2 種類の細胞で構成されています。 グリア細胞はニューロンに密接に隣接しており、サポート、栄養、分泌、保護の機能を果たします。
ニューロンは、神経組織の基本的な構造および機能単位です。 その主な特徴は、神経インパルスを生成し、他のニューロンや作動器官の筋細胞や腺細胞に興奮を伝達する能力です。 ニューロンは、本体とプロセスで構成されます。 神経細胞は、神経インパルスを伝達するように設計されています。 表面のある部分で情報を受け取ると、ニューロンはそれを表面の別の部分に非常に素早く送信します。 ニューロンのプロセスは非常に長いため、情報は長距離に渡って伝達されます。 ほとんどのニューロンには 2 種類の突起があります。短くて太く、体の近くで分岐する樹状突起と、長く (最大 1.5 m) 薄くて最後でのみ分岐する軸索です。 軸索は神経線維を形成します。
神経インパルスは、神経線維に沿って高速で伝わる電波です。
実行される機能と構造的特徴に応じて、すべての神経細胞は、感覚神経細胞、運動神経細胞(実行神経細胞)、および介在神経細胞の 3 つのタイプに分類されます。 神経の一部として走る運動線維は筋肉や腺に信号を伝え、感覚線維は臓器の状態に関する情報を中枢神経系に伝えます。
これで、受け取ったすべての情報をテーブルに結合できるようになりました。
生地の種類(表)
|
ファブリックグループ |
生地の種類 |
組織構造 |
位置 |
|
| 上皮 | フラット | 細胞の表面は滑らかです。 細胞は互いにしっかりと隣接している | 皮膚表面、口腔、食道、肺胞、ネフロン被膜 | 外皮、保護、排泄(ガス交換、尿排泄) |
| 腺状 | 腺細胞は分泌物を生成します | 皮膚腺、胃、腸、内分泌腺、唾液腺 | 排泄(汗、涙の分泌)、分泌(唾液、胃液、腸液、ホルモンの生成) | |
| 繊毛虫(繊毛虫) | 多数の毛を持つ細胞(繊毛)で構成されています | 航空会社 | 保護(繊毛が塵粒子を捕らえて除去する) | |
| 接続詞 | 緻密な繊維質 | 細胞間物質のない、繊維状で密に詰まった細胞のグループ | 皮膚そのもの、腱、靱帯、血管の膜、目の角膜 | 外皮、保護、運動 |
| 緩い繊維質 | 緩く配列された線維細胞が互いに絡み合っています。 細胞間物質は構造を持たない | 皮下脂肪組織、心膜嚢、神経系経路 | 皮膚を筋肉に接続し、体内の臓器をサポートし、臓器間の隙間を埋めます。 体の体温調節を提供します | |
| 軟骨性 | カプセルの中に円形または楕円形の細胞が存在し、細胞間物質は緻密で弾力性があり、透明です。 | 椎間板、喉頭軟骨、気管、耳介、関節面 | 骨の擦れる表面を滑らかにします。 気道と耳の変形に対する保護 | |
| 骨 | 長いプロセス、相互接続された細胞間物質 - 無機塩およびオセインタンパク質を持つ生きた細胞 | 骨格 | サポート、モーター、保護 | |
| 血液とリンパ | 液体結合組織は、形成要素(細胞)と血漿(有機物質と無機物質が溶解した液体、血清とフィブリノーゲンタンパク質)で構成されています。 | 全身の循環系 | O2と栄養素を体全体に運びます。 CO 2 と異化生成物を収集します。 身体の内部環境、化学物質、ガス組成の一定性を確保します。 防御(免疫)。 規制(体液的) | |
| 筋肉質 | クロスストライプ | 長さ10cmまでの多核円筒形細胞で、横縞模様がある。 | 骨格筋、心筋 | 身体とその部分の自発的な動き、顔の表情、会話。 心室に血液を押し出すための心筋の不随意(自動)収縮。 興奮性と収縮性の特性がある |
| スムーズ | 先端が尖った長さ 0.5 mm までの単核細胞 | 消化管の壁、血管およびリンパ管、皮膚の筋肉 | 内部中空器官の壁の不随意収縮。 皮膚に毛を生やす | |
| 神経質 | 神経細胞(ニューロン) | さまざまな形状とサイズの神経細胞体(直径 0.1 mm まで) | 脳と脊髄の灰白質を形成します | より高い神経活動。 身体とのつながりは、 外部環境。 条件反射と無条件反射の中枢。 神経組織には興奮性と伝導性の特性があります |
| ニューロンの短いプロセス - 樹状突起の枝分かれ | 隣接するセルのプロセスと接続する | あるニューロンの興奮を別のニューロンに伝達し、体のすべての器官間の接続を確立します。 | ||
| 神経線維 - 軸索(神経突起) - 長さ1.5メートルまでのニューロンの長い突起。 臓器は分岐した神経終末で終わる | 体のすべての器官を支配する末梢神経系の神経 | 神経系の経路。 それらは、遠心ニューロンを介して神経細胞から末梢に興奮を伝達します。 受容体(神経支配器官)から求心性ニューロンに沿って神経細胞まで。 介在ニューロンは、求心性 (感受性) ニューロンから遠心性 (運動) ニューロンに興奮を伝達します。 |
1) トロフィー機能。
表層結合組織はすべての血管を覆っているため、血液と他の組織の間の物質の交換は結合組織の義務的な参加によって起こります。本質的に、緩い結合組織は血液と他の組織の間の物質の交換を調節します。
2) サポート機能。
サポート機能には、実質と形状形成の 2 種類があります。
— 間質機能。
緩い結合組織は、内臓の骨格である間質を形成します。
- 形状形成。
密な結合組織は臓器の形状を形成する臓器カプセルを形成します。
3) 保護機能。
結合組織は、機械的保護ではなく、主に免疫保護の機能を果たします。 免疫防御は、マクロファージ、マスト細胞、および結合組織によって産生される抗体によって行われます。 ただし、密な結合組織は機械的保護として機能します。
4) 機械的機能とは、動きを組織化する機能です。
高密度の結合組織は、筋骨格系の組織に関与する腱と靱帯を形成します。
5) プラスチック機能。
再生組織への疎性結合組織の参加。 生命活動の過程で臓器に欠陥や傷が形成されると、それらは疎性結合組織で満たされます。 その結果、臓器の形状が復元されます。
結合組織の機能の調節は、細胞、器官、生物のレベルなど、組織のあらゆるレベルで行われます。 細胞レベルでは、細胞膜と密接に関連するエフェクター物質、および細胞間空間に放出されるメディエーター:リンホカイン、モノカイン、フィブロカイン、ラブロカイン(それぞれ、リンパ球、単球、線維芽細胞、組織好塩基球のメディエーター)を介した細胞間の接触が重要です。 細胞膜上に対応する受容体が存在する特異的メディエーターに加えて、プロスタグランジン、ムラミダーゼ、フィブロネクチン、プロテアーゼなどの非特異的メディエーターもあります。
結合組織の要素間の関係はフィードバックの原理に従って実行され、通常の状態では応答の適切性が保証され、病理学では高い適応性と信頼性が保証されます。 細胞間の協力的な相互作用に基づく「ダウン」自己調節は、階層的な「トップダウン」原理に基づいて構築された内分泌および神経調節によって補完されます。
この点で、重要な役割は下垂体前葉のホルモンであるソマトトロピンに属します。 それは、結合組織細胞の増殖とその中の合成プロセスを刺激します。 同時に、コルチコトロピンとグリココルチコイドは増殖を阻害し、線維芽細胞の早期分化と成熟を引き起こし、これにはコラーゲン生成の障害が伴います。 結合組織の構造と機能の調節におけるインスリンの役割は、ヒアルロン酸とコンドロイチン硫酸の交換を促進することです。 どうやら、これは、結合組織全般、特に糖尿病(糖尿病性血管障害)における血管壁の重篤な障害を説明しているようです。
結合組織系の調節不全も、あらゆるレベルで発生する可能性があります。 結合組織内では、確立された自己調節関係およびその実質細胞との関係が破壊される可能性があります。 この障害は、主に 1 つまたは別の臓器 (リウマチでは関節、全身性エリテマトーデスでは皮膚) に現れます。 最後に、結合組織の調節不全は、すべての臓器および体全体に現れる可能性があります(先端巨大症に伴う骨の過剰成長、小人症、甲状腺機能低下症に伴う粘液性浮腫)。
結合組織の状態は老化において重要な役割を果たします。 かつて A.A. ボゴモレツは、「老化はまさに結合組織から始まる」と指摘しました。 彼は、老化の理由を、その高分子成分に物理的および化学的変化が起こるという事実、つまり「細胞のコロイドやミセロイドが成熟し、それらが沈殿物や凝集物に変化し、細胞の生命活動を阻害する生物学的に不活性な封入体を形成する」ことにあると考えました。
現代の研究では、年齢とともに実際に結合組織に変化が起こり、その栄養機能、保護機能、その他の機能が破壊されることがわかっています。 皮膚、腱、軟骨組織、および大動脈では、細胞の数、細胞の大きさ、核の大きさが減少します。 たとえば、老人の炎症時には線維芽細胞の再生が遅くなります。 線維構造では、加齢に伴う変化はコラーゲン線維間の架橋数の増加によって表されます。 過剰な量の高分子「架橋」を持つコラーゲンは、新たな特性を獲得します。 温度の影響に対する耐性が高まり、溶解性と水と結合する能力が低下します。 通常は代謝的に安定していますが、コラーゲンはますます不活性になります。 その自己再生が遅くなり、必然的に分子内に新たなエラーが蓄積することになります。 主要物質に含まれるヒアルロン酸の量は減少します。これはおそらく年齢とともに体の水分と結合する能力を低下させます。 同時に、血管壁内のコンドロイチン硫酸の量が増加します。 後者は、硫酸化グリコサミノグリカンがカルシウムイオンに対する親和性を有するため、血管の石灰化を促進します。
結合組織の保護機能は、皮膚、粘膜、線維性被膜の構造、および特殊な関門(血液脳関門のグリア)の構造に関与することによって、機械的障壁の作成。 結合組織細胞は、食作用 (マクロ食細胞) などの保護の形態、および損傷した組織を正常な組織 (肉芽シャフト) から区切る能力を決定します。 最後に、結合組織細胞はリンパ球と相互作用して免疫応答に関与します。
A.A. ボゴモレッツは、結合組織の保護的役割を初めて完全に認識しました。 彼と彼の学生らは、結合組織の反応性の低さは、一般に、より重篤な疾患の経過と組み合わされていることを示した。つまり、傷の治りが遅くなり、骨折の治りが悪くなるということである。
結合組織の機能
これらすべてのことから、結合組織に影響を与えることによって病気の経過を悪化または改善し、重症度を下げることが可能であると考えることができました。 その機能を刺激する方法を見つけることだけが必要でした。 理学療法、マッサージ、日光浴、食事療法などの方法によって特定の結果を達成することもできますが、A.A. ボゴモレツは、結合組織のすべての要素が体内のどこに存在していても、それらの要素を特異的に選択的に刺激することを念頭に置いていました。 その結果、これは免疫学的に達成されました。 結合組織が豊富な臓器(骨髄、脾臓)を備えた動物(馬)の免疫化の結果得られる血清を人に注射すること。 この血清は ACS (抗網状細胞傷害性血清) と略称され、A. A. ボゴモレッツと彼の学生によって詳細に研究され、その後臨床で使用されました。
抗網様細胞傷害性血清の効果はその用量に依存することが判明した。 大量の血清には細胞毒性作用がありました。 結合組織細胞を破壊し、小さい - 刺激します。 これは、組織のマクロ食細胞要素の代謝の増加と食作用の活性化、抗菌抗体の力価の増加、水電解質と脂肪の代謝の正常化によって表されました。 酵素プロセスの活性化により、結合組織は、さまざまな代謝疾患や老化中に蓄積するバラスト物質から解放されます。 この場合、体の細胞に非特異的な効果を及ぼし、一般的な刺激効果を構成する物質が形成される可能性があります。
講義検索
骨格の機械的機能
1.サポート機能それは、骨がそれに付着した軟組織(筋肉、筋膜、その他の器官)を支持し、内臓が位置する空洞の壁の形成に関与しているという事実にあります。
2. スプリング機能衝撃や衝撃を和らげる構造(軟骨パッド、接続骨間の関節軟骨など)が骨格に存在するためです。
3. 保護機能それは、骨格が重要な器官の入れ物を形成し、外部の影響からそれらを保護するということです。
4. モーター機能これは、可動関節によって接続され、神経系によって制御される筋肉によって駆動される、長短のレバーの形をした骨の構造によって可能になります。
5.反重力機能それは、骨格が地面から上がった体の安定性をサポートしているという事実に現れています。
さらに、骨は血管、神経、筋肉の走行方向、体の形や大きさを決定します。
骨格の生物学的機能
1. 代謝機能- 骨格は代謝(特にミネラル代謝)に関与しており、リン、カルシウム、鉄などのミネラル塩の貯蔵庫となっています。
2. 造血機能骨の内部には、中央の造血器官である骨の有機部分である赤い骨髄があるという事実によるものです。
3. 免疫機能赤色骨髄にも関連しています。赤色骨髄には自立した造血幹細胞の集団が含まれており、そこから免疫系細胞やリンパ球も形成されます。
臓器としての骨
各管状骨には次の部分があります。
1. 骨幹(骨体)は、成人の黄色の骨髄を含む骨管で、それぞれ支持と保護の機能を果たします。
2. 骨幹端中骨端軟骨に隣接する骨幹(骨幹の端)は、骨幹とともに発達しますが、骨の長さの成長に関与し、海綿状物質で構成されています。
3. 骨端(各管状骨の関節端)は中骨端軟骨の反対側に位置します。
4. 先端部(骨端近くにある骨の突起)。
骨の分類
成人の人間の骨格を構成する個々の骨の数は 200 以上 (206 個の骨) です。 骨は大きさや形が異なり、体の特定の位置を占めています。 外形に基づいて、骨は長いもの、短いもの、広いもの、および混合されたものに区別されます。
しかし、解剖学的分類の基礎となる 3 つの原則、つまり形態(構造)、機能、発達に基づいて骨を区別する方がより正確です。 この観点から、次の骨のグループが区別されます。
![]()
![]() 骨格
骨格
管状海綿状平らな混合空中浮遊
長い長い短い頭蓋骨
短い種子骨ベルト
骨の接続
ボーンの接続には 3 つのタイプがあります。
1) 継続的な接続 (関節症)、骨の間に結合組織または軟骨の層がある場合。 接続している骨の間に隙間や空洞はありません。
2) 断続的な接続 または関節(可動性関節症、または滑膜関節) - 骨と関節包の内側を覆う滑膜の間に空洞がある場合。
3) ハーフジョイント または結合(半関節症)、結合している骨の間の軟骨または結合組織層に小さな隙間がある場合。
1. 継続的な接続 - 関節症。 骨を接続する組織の構造に応じて、これらの接続は次のグループに区別されます。
— 線維性(結合組織)または結合組織。
— 軟骨性(軟骨結合症);
- 骨の接続(癒合症)。
- 弾性;
- 筋肉のつながり。
線維性関節(結合症) これらは、高密度の線維性結合組織を介した強力な接続です。 これらには次のものが含まれます。
A) 膜または骨間膜.
b) バンドル
V) 縫い目:
- 鋸歯状(前頭骨と頭頂骨の接続など)。
- 鱗状(たとえば、側頭骨と頭頂骨の接続)。
- 滑らか (顔の頭蓋骨間の接続など)/
G) ハンマーで叩く
軟骨関節(関節症) それらは骨と軟骨の間の接続です。 存在期間に応じて、関節症は次のようになります。
A) 一時的– 一定の年齢まで存在し、その後は癒合症によって置き換えられます(たとえば、骨盤帯の骨の間)。
b) 永続– ある年齢まで存在し、その後、癒合症によって置き換えられます(たとえば、側頭骨の錐体と隣接する骨盤帯の骨の間)。
伸縮性のある接続 結合組織や繊維状化合物のような強度はありません。
骨の関節 (癒合症): 骨の間の空間では、結合組織が骨に変化するか、最初に軟骨に変化し、次に骨に変化します。
筋肉のつながり それらは、横紋筋の助けを借りて、2つ以上の骨を接続し、可動で長さを変えることができます。
2. 不連続な関節または関節 (可動性関節症)最も高度なタイプの骨接続です。
各関節では次のことが区別されます。 必須の要素:
- 関節表面は軟骨で覆われています。
- 関節包または滑液包;
- 少量の関節液を含む関節腔。
一部の関節には、関節円板、半月板、および関節唇の形の補助構造もあります。
関節面 ほとんどの場合、骨の関節において互いに対応しています。
結合組織 - 構造、機能、構成
それらは関節軟骨で覆われており、関節表面の滑りを促進し、衝撃を和らげます。
関節包 関節面の端に沿って、または関節面からわずかに後退して関節骨まで成長し、関節腔を密閉して取り囲みます。
カプセルには、外側の繊維層と内側の滑膜層の 2 つの層があります。
繊維層場所によっては靱帯を形成します。これが肥厚してカプセルを強化し、受動的ブレーキとしても機能し、関節の動きを制限します。
滑膜層薄い。 それは内側から線維層を覆い、関節軟骨で覆われずに骨の表面に続きます。
関節腔 それは関節面と滑膜によって制限された密閉されたスリット状の空間です。 関節腔には少量の滑液が含まれています。
3. 半関節または結合(半関節症) -接続を連続から不連続に、またはその逆に移行します。 これらは軟骨性または繊維状の化合物であり、その厚さには隙間の形の小さな空洞があります。
関節の分類
関節では、関節面の構造 (形状、曲率、サイズ) に応じて、異なる軸の周りで動きが発生することがあります。 関節の生体力学では、次の回転軸が区別されます: 1) 正面、2) 矢状、3) 垂直。 さらに、円運動も区別されます。
ジョイントは次の基準に従って分類されます。
- 関節面の数による。
- 関節面の形状に応じて;
- 機能別。
私。関節面の数に基づいて、次のように区別されます。
A) 単純なジョイント– 2 つの関節面があります (例: 上腕骨、指節間)
b) 複雑な関節– 関節面が 2 つ以上ある (肘、膝など)。 複雑なジョイントは、動作を個別に実行できるいくつかの単純なジョイントで構成されます。
V) 複雑な関節– 関節包の内側に関節内軟骨が含まれており、関節を 2 つの部屋に分割します (例、顎関節、膝)。
G) 結合ジョイント– 互いに分離された複数の関節の組み合わせを表します。関節は互いに離れて配置されていますが、一緒に機能します(たとえば、両方の顎関節、近位橈尺関節と遠位橈尺関節など)。
II.形状と機能に従って、次のように分類が行われます。関節の機能は、動作が行われる軸の数によって決まります。 これらの軸の数は、関節の関節面の形状によって異なります。 これに基づいて、ジョイントは次のように区別されます。
1. 一軸ジョイント(円筒状または回転式でブロック状):
2.二軸ジョイント(楕円体、サドル、顆):
3. 三軸または多軸ジョイント(球形、ナット形、平ら):
©2015-2018 poisk-ru.ru
動物の結合組織
動物の体重の大部分は結合組織によって形成されます。 それらは軟骨、骨、腱、靭帯で構成されています。
結合組織の特徴
結合組織の構造は、動物が異なれば、また同じ生物の部位が異なれば異なります。 さらに、それらの構造の共通の特徴は、細胞が細胞間物質の塊の中に点在しているように見えることです。 結合組織には、異なる機能を実行するいくつかの種類があります。
線維性結合組織
線維性結合組織は動物の体のいたるところに見られます。 皮膚と筋肉を接続し、望ましい位置に保持し、臓器を接続します。 このタイプの組織の細胞は、細胞間物質を形成する高密度の繊維ネットワークに囲まれています。
骨
骨組織は骨格の骨、つまり脊椎動物の内部支持体を形成します。 骨組織は、骨に強度を与えるミネラル物質と、弾力性を与える有機物質で構成されています。
結合組織の構造と機能、主な細胞の種類
この組み合わせは、骨組織がサポート機能を実行するのに役立ちます。
骨細胞は動物の生涯を通じて生き続け、細胞間物質を分泌します。 細胞は、骨間物質に存在する多数の突起によって互いに接続されています。
骨組織は骨を形成します。 骨組織によって形成される骨の成長と栄養は、骨を覆う骨膜によって確保されています。
軟骨組織
軟骨組織は骨の頭を覆い、関節にあり、骨格に柔軟性を与えます。
軟骨組織の細胞は、単独またはグループで、弾性のある細胞間物質の中に浸されています。 サメやエイの骨格には骨組織はなく、完全に軟骨でできています。 人間では、耳介と鼻の先端で軟骨を感じることができます。
血
血液は特別な結合組織です。 液体の細胞間物質である血漿が含まれています。 血漿には、赤血球 (赤血球)、白血球 (白血球)、血小板 (円形、卵形の細胞、または血小板) という血液細胞が含まれています。
血液が最も小さな血管である毛細血管を通って移動すると、溶解した状態の栄養素が細胞間隙に浸透します。 その結果、組織液が形成されます。 そこからリンパ液(リンパ液、つまり水分、きれいな水)が発生し、リンパ管に集まり、そこから再び血液に入ります。
血液、リンパ液、組織液は体の内部環境を作り出します。

脂肪組織
脂肪組織も結合組織です。 多数の脂肪細胞から構成されています。 この組織は主に皮下脂肪層に存在します。 栄養が不足した場合に体が利用できる脂肪を蓄えます。 さらに、脂肪組織は動物の熱を保持し、外部の衝撃から保護するのに役立ちます。
講義検索
結合組織。 体内の位置、種類、構造、機能。
細胞と大量の細胞間物質で構成されています。 細胞間物質には繊維と基質が含まれます。 繊維は強度と弾力性を提供します。
繊維は次のように分類されます。
û コラーゲン
û 網状
û弾性
コラーゲン繊維にはコラーゲンというタンパク質が含まれており、耐久性に優れています。
網状線維は赤骨髄、リンパ節、脾臓の一部です。 薄くて緻密なネットワークを形成できます。
弾性繊維にはエラスチンというタンパク質が含まれていますが、コラーゲンよりも耐久性が低く、簡単に伸びてしまいます。
細胞間に属する主な物質は、細胞と繊維の間の空間を満たします。
機能はさまざまです。
- 支持結合組織は、骨格の骨、軟骨、靱帯、腱、筋膜の一部です。 支持機能は、高密度の繊維組織(靱帯および腱)、骨および軟骨組織によって実行されます。
- 栄養 - この機能は血液とリンパによって行われます(他の組織に栄養素を提供します)。
- 機械的結合組織は、柔らかい骨格、つまり間質の形成に関与します。
- 結合組織は造血、つまり造血に関与しています。
- 結合組織は食作用に関与しています。
- 結合組織は再生に関与しています。
- 呼吸機能 - 組織や器官で起こるガス交換のプロセスに関与します。
結合組織には結合組織自体が含まれ、疎な繊維と密な繊維が含まれます。 骨格結合組織(軟骨および骨)、および特別な特性を持つ結合組織(脂肪組織、血液、リンパおよび造血組織)。
疎性線維性結合組織 (FCT)。
PBST は臓器間の空間を埋めます。
PBST には次のセルが含まれます。
線維芽細胞は平らな紡錘形の細胞です。
結合組織の機能
創傷治癒と瘢痕組織の形成に関与します。
ü マクロファージは、異物を捕らえて消化する細胞です。
ü マスト細胞は血液凝固を防ぐヘパリンを生成します。
ü 血漿 - 抗体の合成に関与します。
抗体は感染を防ぐタンパク質です。
ü 脂肪細胞 - 予備脂肪を蓄積する能力があります。
ü 色素細胞 - メラニン色素の粒子が含まれています。
高密度線維性結合組織 (DFCT)。
この生地は繊維が緻密に配列されています。 細胞間物質はほとんどありません。 PVST は、靱帯、腱、筋膜、膜の一部です。
筋膜は、筋肉が配置されている薄い結合組織の鞘です。
コラーゲン線維が多く含まれています。
軟骨組織は、軟骨細胞と緻密な細胞間物質から構成されています。
細胞間物質にはさまざまな繊維が存在します。
ヒアリン
弾力性のある
- 繊維質
硝子軟骨は肋骨の一部です。 肋骨と胸骨の接合部に位置します。
弾性軟骨は耳介と喉頭の軟骨の一部です。 カルシウムは弾性軟骨には決して沈着しません。
線維軟骨は椎間板を形成し、下顎関節を覆っています。
骨。
細胞と細胞間物質から構成されています。
細胞間物質には主物質が含まれており、無機塩(カルシウム、マグネシウム)が多く含まれています。
有機物質 - 脂肪、タンパク質、炭素を含む炭水化物。
無機物質 - 無機塩。
そのおかげで骨が丈夫になります。 骨にはカルシウム塩が多く含まれています。 カルシウム塩が不足すると発症する 骨粗鬆症 . 骨がもろくなり、骨折する可能性があります。
骨に含まれる有機塩の中で最も多いのは、 オセナ骨に柔軟性を与えます。
骨では、破壊と新しい細胞の形成のプロセスが常に発生します。
骨細胞には 3 種類あります。
- 骨芽細胞は、骨組織を形成する細胞です。
- 骨細胞は、骨芽細胞から形成される細胞です。
- 破骨細胞は、骨組織を破壊する細胞です。
骨組織には 2 種類あります。
Ø 粗い繊維
Øプレート
頭蓋骨の縫合部には粗い繊維組織が見られます。 コラーゲン線維と骨細胞で構成されています。
層状組織は粗い繊維状組織よりも密度が高く、すべての骨は層状組織から構築されます。 また、板状のコラーゲン線維や細胞も多数含まれています。
骨の機能単位は、 オステオン。
脂肪組織
これは結合組織であり、その大部分は脂肪細胞、つまり脂肪細胞によって占められています。 2つのタイプがあります:白色脂肪組織(表面および深部の蓄積を形成する)、褐色脂肪組織(肩甲骨の間、脇の下、首の大きな血管の領域に位置する)。
血液とリンパ
それらは液体部分と形成された要素で構成されます。
©2015-2018 poisk-ru.ru
すべての権利はその作者に帰属します。 このサイトは著作者であることを主張しませんが、自由に使用できます。
著作権侵害および個人データ違反
結合組織は体内に広く分布しています。 それは、内臓、皮膚、靱帯、腱、筋肉および神経の膜、血管壁に見られます。
結合組織は、線維芽細胞、組織球、大食細胞、組織好塩基球、および細胞間物質(線維 - コラーゲン、弾性物質および基質)を含む細胞で構成されています。
組織や器官には、さまざまな形成段階の細胞があります。
さまざまな種類の細胞と細胞間物質の組み合わせによって、結合組織の多種多様な構造と機能が決まります。
結合組織の機能 :
1. 栄養(細胞栄養 - 血液、リンパ)
2. 防御 – (貪食、抗体形成)
3. 形状形成(臓器実質、筋膜の形成)
4. 再生(組織再生、創傷治癒)
5.排泄
結合組織細胞と細胞間物質.
線維芽細胞は平らな紡錘形の細胞であり、結合組織の主要な細胞であり、運動性があります。
1. 細胞間物質の形成、繊維構造の合成:
コラーゲン、エラスチン、レチクリン
2.分割可能
3. それらは未分化型から発生し、他の細胞に変化する可能性があります
4. 創傷治癒と瘢痕組織の形成に参加します。
肝臓マクロファージ – 星細胞
肺マクロファージ - 肺胞食細胞
漿液腔のマクロファージ - 胸膜および腹膜のマクロファージ
骨組織 - 骨芽細胞
神経質 - ミクログリア細胞。
大食細胞 - 主な機能 - 食作用 - 有害な異物や異物、死んだ細胞、細菌、ウイルスなどを体から除去します。
組織好塩基球(肥満細胞) – ヘパリン、ヒスタミン、セロトニンを生成します。
形質細胞– 抗体を生成します – 緩い結合組織、腸粘膜、大網、リンパ節、骨髄に存在します。
脂肪細胞– 予備脂肪を蓄積します。 脂肪細胞が蓄積すると脂肪組織が形成されます。
網様細胞– メッシュを形成し、そのループ内に主要組織の細胞が配置されます。
外膜細胞– 血管壁および中空器官の外層に位置します。
色素細胞– メラニン粒子を含み合成し、結合組織、肛門周囲の皮膚、乳腺の陰嚢と乳輪の皮膚、眼の脈絡膜に見られます。
結合組織の細胞間物質 .
1/ ムコ多糖生体高分子 B および U - ゼリー状物質 (ヘパリン、ヒアルロン酸) - ネットワークと細孔を形成
2/ 糖タンパク質ポリマー B および U – 線維芽細胞で形成されます。 弾力のあるコラーゲン線維を形成します。
3/繊維:
コラーゲン– 臓器や組織の特定の構造をサポートします
弾性のある- ワイドループネットワークの形式で。
網状- ネットワークを形成し、リンパ節、脾臓、骨髄などの一部です。
7. 栄養性(適切な結合)組織をサポートします。
ルースファイバーST- 血管の一部であり、リンパ系形成の基礎を形成します。
細胞: 線維芽細胞
細胞間物質がたくさんある
繊維: コラーゲンと弾性 – 拡散して絡み合って位置します。
緻密な繊維状のST。
繊維は互いに平行に配置され、束に集められます。
細胞は線維芽細胞です(数はほとんどありません)。 生地は耐久性があり、柔軟性があり、伸びません。 その繊維は負荷がかかった状態で張力線と平行に配置されます。
強膜、角膜、腎被膜、髄膜に含まれています。
緻密な繊維状の未形成ST。
繊維は互いにしっかりと隣接しており、絡み合っています
細胞や基質がほとんどありません。 この組織は次のもので構成されています: - 靭帯
動脈壁
特別な特性を持つ結合組織- 網様組織
構成: 細胞 - 網状赤血球 - は線維芽細胞、マクロファージに変わることができます。
線維は網状で、複雑な神経叢を形成しています。
RT は骨髄および骨髄組織の基礎です。
網状組織は扁桃腺の一部であり、腸粘膜を形成します。
軟骨組織 – 骨組織とともに結合組織をサポートすることを指します。
軟骨の構造:
1. 細胞– 軟骨芽細胞と軟骨細胞 – 単独またはグループで存在する
2. 細胞間物質:
a) 主要物質が高密度である
b) 繊維 – コラーゲン (さらに多くあります)
弾性のある
軟骨の種類(細胞間物質の構造による)
1/ 硝子軟骨組織(硝子体軟骨) – より多くの塩基性物質が含まれています。 HCT は以下で構成されます: - 胎児骨格
骨の関節面
肋骨の軟骨部分
2/ エラスティックXT– ヒアリンから形成されます。 弾力性のある繊維がより多く含まれています。 弾性軟骨 - 耳介の基礎、喉頭の軟骨、外耳道の壁。 透明度は低く、色は黄色がかっていますが、形状を復元することができます。
3/ ファイバースXT– コラーゲン線維は束に集められ、順序付けされます。
VXT は GC よりも強力ですが、柔軟性に劣ります。 恥骨結合と椎間板を形成します。
線維性軟骨組織は衝撃吸収材として機能します。
軟骨は補助的な機械的機能を果たします。
骨 - 支持結合組織は、軟骨組織とともに骨格の骨を形成します。
骨組織の構造:
骨芽細胞 – 骨組織の主要物質を形成し、骨形成ゾーン(成長中の骨の表面)に位置します。
破骨細胞 - 突起のある大きな多核細胞。 骨と軟骨の破壊に関与し、湾または裂孔が形成されます(骨は酵素のおかげで吸収されます)。
骨細胞 – 骨芽細胞から形成されます。 芽が出ています。 彼らの体は骨空洞の中にあり、その突起は骨小管に入ります。
細胞間物質は石化されています。
コラーゲン線維(オセイン線維) – 骨に柔軟性と可塑性を与えます。
骨組織の種類:
1. 粗い繊維質- 胎児およびヒトの頭蓋骨の縫合部および骨への腱の付着箇所。
粗い骨組織内のオセインコラーゲン線維は束を形成し、その間に骨細胞が骨腔内にあります。
2. ラメラ(細繊維) - 骨格のすべての骨。
コラーゲン線維は、プレート内またはプレート間に平行な束となって配置されています。
3. 象牙質– 象牙芽細胞は象牙質(体)の外側にあり、そのプロセスは象牙質内の尿細管を通過します。
層状組織の形態
1. 緻密な骨質
2. 海綿骨物質それらは骨を構成します
緻密な骨物質では、プレートが特別な順序で配置され、骨密度(骨幹)が与えられます。
海綿状の骨物質では、プレートは横棒(骨端、短骨)を形成します。
緻密な骨物質では、骨プレートは一種の管状システムであるオステオン(骨の構造単位)を形成します。
骨プレートは、血管が通過するハバース管(骨骨の中心にある空洞)の周囲に同心円状に配置されています。
骨細胞は骨板の間に位置します。
骨膜(骨膜)– 2 つの層からなる結合組織膜。
外層はより高密度の結合組織でできており、筋肉の腱や靭帯がそれに付着しています。
内層はコラーゲンと弾性線維、骨芽細胞、
破骨細胞。
骨の成長中、骨芽細胞は骨の形成に関与します。 骨膜には、骨を貫通して骨に栄養を与える多数の血管と神経が含まれています。 骨折における骨の再生は、骨膜によって起こります。骨膜は、骨折部位を覆って成長し、折れた骨の端を接続し、その周囲に骨組織の塊である仮骨を形成します。
エンドスト– 髄管の側から骨を覆う膜。
筋。
古代人は「人生は動いている」と言いました。
あなたは頭を向け、瞬きし、息を吸い、遠くを見つめ、何かを言いました。 毎分、体内では何千もの筋線維と細胞が収縮します。 これに加えて、心臓は鼓動し、胃は鳴り、尿管は尿を腎臓から膀胱に穏やかに運び、血管は常に一定の血圧を維持します。
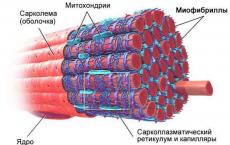
人体の運動プロセスは、収縮性という特定の特性を持つ筋肉組織の収縮によって引き起こされます。
組織学者が強調する 3種類の筋肉組織:
1. 平滑筋組織。
2. 縞模様の骨格筋組織。
3. 横紋心筋組織。
第8章 結合組織
第8章 結合組織
結合組織は、 大人数のグループ間葉由来の組織は、原則として多種であり、細胞間物質(繊維構造と非晶質成分)が優勢であり、体の内部環境の組成の一定性の維持に関与しています。
結合組織は人間の体重の 50% 以上を占めます。 それは、臓器の間質、他の組織の間の層、皮膚の真皮、および骨格の形成に関与します。 結合組織の多機能性は、その組成と組織の複雑さによって決まります。
結合組織の機能。結合組織は、栄養、保護、支持(生体力学的)、可塑性、形態形成など、さまざまな機能を果たします。 広義の栄養機能は、さまざまな組織構造の栄養の調節、代謝への参加、および身体の内部環境の組成の一定性の維持に関連しています。 この機能の実現において、主な役割は、水、塩分、栄養素分子の輸送が行われる主要な物質である統合緩衝媒体によって演じられます。 保護機能は、非生理的・機械的影響(損傷)から身体を保護し、外部から侵入する異物や体内で形成される異物を中和することです。 これは、物理的保護(骨組織)、ならびに細胞性および体液性免疫の反応に関与するマクロファージおよび免疫担当細胞の貪食活性によって提供されます。 サポート(生体力学的)機能は、主に、すべての器官の繊維基盤を形成するコラーゲンと弾性線維、および骨格組織の細胞間物質の組成と物理化学的性質(石灰化)によって提供されます。 細胞間物質の密度が高くなるほど、それを支える生体力学的機能がより重要になります。 結合組織の可塑的機能は、変化する生活条件への適応、再生、および臓器が損傷した場合の欠損の置換への参加において発現されます。 形態形成
(構造形成)機能は、組織複合体の形成と臓器の一般的な構造組織の確保(カプセルの形成、臓器内隔壁)、さまざまな組織の細胞の増殖と分化に対するその成分の一部の制御影響によって現れます。
結合組織の分類。結合組織の種類は、細胞の差異、線維、および組織の組成と比率が異なります。 物理的及び化学的性質非晶質の細胞間物質。 結合組織は、結合組織そのもの(疎性結合組織と特殊な性質をもつ結合組織)と骨格組織に分けられます。 後者は、3 種類の軟骨組織 (硝子質、弾性、線維性)、2 種類の骨組織 (網状線維質および層状)、さらに歯のセメントと象牙質に分けられます (図 8.1)。
結合組織の組織形成。結合組織の発達の源は間葉です(ギリシャ語から。 メソス- 平均、 エンチマ- 充填質量)。 これは胚の原始(いくつかの考えによれば、胚組織)の1つであり、中間胚葉の緩んだ部分です。 間葉の細胞要素は、皮分節、強膜分化、内臓層、および内臓層の分化の過程で形成されます。 さらに、神経堤(神経節板)から発生する外胚葉間葉(神経間葉)もあります。 胚が発生するにつれて、他の胚細胞とは異なる起源の細胞が間葉に移動します。
スキーム8.1。結合組織の分類
原始的なもの、例えば、神経芽細胞性ディファロン細胞、骨格筋原基由来の筋芽細胞、色素細胞など。
したがって、胚の発生の特定の段階から、間葉はさまざまな胚葉および胚組織の基本から生じる細胞のモザイクになります。 しかし、形態学的には、すべての間葉細胞は互いにほとんど違いがなく、間葉内の異なる性質の細胞を明らかにするには、非常に感度の高い研究方法(通常、免疫細胞化学、電子顕微鏡)のみが必要です。 間葉は人間の発達の胎児期にのみ存在します。 出生後、低分化(多能性)細胞のみが疎性線維性結合組織(外膜細胞)の一部として人体に残り、特定の組織系内でさまざまな方向に分岐して分化することができます。
結合組織の組織形成には胚期および胚後の組織形成があります。 胚の組織形成の過程で、間葉は他の組織が形成される前に組織構造の特徴を獲得します。 このプロセスは、異なる器官や系で異なる方法で起こり、胚形成の異なる段階での異なる生理学的重要性に依存します。 間葉の分化では、胚と胚外器官の両方でトポグラフィーの非同期性、高い細胞複製率と線維形成が観察されます。 通常の生理学的条件下での胚後組織形成はよりゆっくりと起こり、組織の恒常性の維持、低分化細胞の増殖、および死にかけている細胞の置換を目的としています。 これらのプロセスにおける重要な役割は、細胞間の組織内相互作用、誘導因子および阻害因子(インテグリン、細胞間接着因子、機能的負荷、ホルモン、酸素化、低分化細胞の存在)によって演じられます。
結合組織組織の一般原則。結合組織の主な構成要素は、細胞派生物です。コラーゲンと弾性タイプの線維構造、統合的な緩衝代謝環境の役割を果たす塩基性(非晶質)物質、および組成の量的および質的比率を作成および維持する細胞要素です。非細胞成分の。
結合組織の細胞要素の器官特異性は、量、形状、比率で表されます。 さまざまな種類臓器の機能に最適に適応した細胞、その代謝と機能。 細胞要素の特異性は、それらの相互作用(個別に位置する細胞結合)、それらの要素の特徴によっても明らかにされます。 内部構造(細胞小器官の組成、核構造、酵素の存在など)。 結合組織の特異性は、体のさまざまな部分にあるさまざまな細胞と非細胞構造の比率にも現れます。 疎な線維性結合組織では、さまざまな異なる細胞と非晶質物質が線維の上に広がっており、密な結合組織では、逆に、その塊の大部分が線維で構成されています。
8.1. 適切な結合組織
8.1.1. 線維性結合組織
緩い結合組織
緩い結合組織 (textus connectivus laxus)血管やリンパ管に随伴し、多くの臓器の間質を形成するため、あらゆる臓器に存在します。 臓器の特徴の存在にもかかわらず、さまざまな臓器の疎性結合組織の構造は類似しています。 それは、さまざまな組織遺伝学的決定と細胞間物質の細胞で構成されています(図8.1)。

米。 8.1.緩い結合組織:
あ- フィルムの準備: 1 - 線維芽細胞。 2 - マクロファージ。 3 - コラーゲン線維。 4 - 弾性繊維。 5 - リンパ球。 b- 線維芽細胞; V- マクロファージ

米。 8.2.分化のさまざまな段階における線維芽細胞の超顕微鏡構造 (N. A. ユリナおよび A. I. ラドスティーナによる、修正あり): 線維芽細胞: あ- 低分化; b- 若い; V- 成熟した; G- 線維細胞。 1 - コア。 2 - ゴルジ複合体。 3 - ミトコンドリア。 4 - リボソームとポリリボソーム。 5 - 顆粒小胞体。 6 - コラーゲン原線維
細胞
結合組織の主な細胞は次のとおりです。 線維芽細胞(原線維形成細胞のファミリー)、 マクロファージ(家族)、 マスト細胞、外膜細胞、形質細胞、周皮細胞、脂肪細胞、そして 白血球、血液から移行する。 時々 顔料
細胞。
線維芽細胞(緯度から。 ファイバ- 繊維、ギリシャ語 爆風- 胚芽、胚芽) - 細胞間物質の成分を合成する細胞:タンパク質(コラーゲン、エラスチン)、プロテオグリカン、糖タンパク質(図8.1、図8.2、8.3を参照)。
間葉系細胞の中には、線維芽細胞の差異を生み出す幹細胞があります:幹細胞、半幹前駆細胞、低分化(不十分に分化)、分化線維芽細胞(成熟、活発に機能)、線維細胞(最終細胞形態) 、筋線維芽細胞と線維芽細胞も同様です。 線維芽細胞の主な機能は、基質と線維の形成、創傷治癒、肉芽組織の発達、異物の周囲の結合組織被膜の形成などに関連しています。形態学的には、このディフェロン内で確認できるのは細胞のみです。 、 で始まります 分化が不十分な線維芽細胞。後者は、円形または楕円形の核と、RNA が豊富な小さな核小体の好塩基性細胞質を備えた小さな細胞です。 セルのサイズは 20 ~ 25 ミクロンを超えません。 これらの細胞の細胞質には、
多数の遊離リボソーム (図 8.2 を参照)。 小胞体とミトコンドリアはあまり発達していません。 ゴルジ複合体は、短い管と小胞のクラスターによって表されます。
分化した成熟線維芽細胞サイズが大きくなり、フィルム上で平らになったものは、40 ~ 50 ミクロン以上に達することがあります (図 8.1 を参照)。 これらは活発に機能している細胞です。 それらの核は軽く、楕円形で、1 ~ 2 個の大きな核小体を含みます。 細胞質は好塩基性で、よく発達した顆粒小胞体を持ち、所々細胞膜と接触しています(図 8.2、図 8.3、 A)。ゴルジ複合体は細胞全体に槽および小胞の形で分布しています。 ミトコンドリアとリソソームは適度に発達しています。
線維芽細胞の細胞質、特に周辺部分には、アクチンやミオシンなどのタンパク質を含む厚さ 5 ~ 6 nm のマイクロフィラメントが存在し、これらの細胞の運動能力を決定します。 線維芽細胞の移動は、線維芽細胞や他の細胞によって合成され、細胞と非細胞構造の接着を提供する糖タンパク質であるフィブロネクチンの助けを借りて、線維芽細胞が支持線維構造(フィブリン、結合組織線維)に結合した後にのみ可能になります。 運動中に線維芽細胞は平らになり、その表面は 10 倍に増加することがあります。
線維芽細胞のプラズマレンマは、さまざまな調節因子の影響を媒介する重要な受容体ゾーンです。 線維芽細胞の活性化には、通常、グリコーゲンの蓄積と加水分解酵素の活性の増加が伴います。 グリコーゲン代謝によって生成されるエネルギーは、細胞によって分泌されるポリペプチドおよびその他の成分を合成するために使用されます。
成熟線維芽細胞におけるコラーゲンおよびエラスチンタンパク質、主要物質および線維の形成に必要なプロテオグリカンの生合成は、特に低酸素濃度の条件下で非常に集中的に行われます。 コラーゲン生合成の刺激因子は、鉄、銅、クロムイオン、アスコルビン酸です。 加水分解酵素の 1 つであるコラゲナーゼは、細胞内の未熟なコラーゲンを分解し、細胞レベルでのコラーゲン分泌の強度を制御していると考えられます。
線維性タンパク質を合成する能力に基づいて、線維芽細胞ファミリーには、造血器官の網様結合組織の網様細胞、結合組織の骨格タイプの軟骨芽細胞および骨芽細胞が含まれます。
線維細胞- 線維芽細胞の発達の決定的な形態。 これらの細胞は翼状の突起を備えた紡錘形です。 それらには、少数の細胞小器官、液胞、脂質、グリコーゲンが含まれています。 線維細胞内のコラーゲンやその他の物質の合成が急激に減少します。
筋線維芽細胞- 形態学的に線維芽細胞に類似しており、コラーゲンを合成する能力だけでなく収縮性も兼ね備えた細胞

米。 8.3.線維芽細胞、筋線維芽細胞、線維芽細胞:
あ-線維芽細胞(A.I.Radostinaによる調製):1 -核。 2 - 顆粒小胞体。 3 - リボソーム。 4 - ゴルジ複合体。 5 - ミトコンドリア。 6 - プラズマモレンマ。 7 - コラーゲン原線維。 b- 筋線維芽細胞 (A.B. シェクターの調製物): 1 - 核。 2 - 顆粒小胞体。 3 - リボソーム。 4 - ゴルジ複合体。 5 - 収縮性フィラメント。 6 - プラズマレンマ
体内タンパク質が大量に含まれています (図 8.3、b を参照)。 線維芽細胞は、平滑筋細胞と機能的に類似した筋線維芽細胞に成長することができることが確立されているが、後者とは異なり、よく発達した小胞体を持っている。 このような細胞が観察されるのは、

米。 8.3.継続:
V- 線維芽細胞(A. B. Shekhter による調製): 1 - コア。 2 - 顆粒小胞体。 3 - リボソーム。 4 - リソソーム。 5 - コラーゲン原線維の断片を含むファゴリソソーム。 電子顕微鏡写真。 倍率 20,000
創傷過程の再生状態および妊娠の進行中の子宮内の肉芽組織。
線維芽細胞- 高い貪食活性と加水分解活性を持つ細胞は、臓器退縮期(たとえば、妊娠終了後の子宮)に細胞間物質の「吸収」に参加します(図8.3、cを参照)。 それらは、原線維形成細胞の構造的特徴(発達した顆粒小胞体、ゴルジ複合体、比較的大きいが少数のミトコンドリア)と、特徴的な加水分解酵素を備えたリソソームを組み合わせています。 それらが細胞外に分泌する酵素の複合体は、コラーゲン線維の接着物質を分解し、その後リソソームの酸性プロテアーゼによるコラーゲンの貪食および細胞内消化が起こります。
マクロファージ(大食細胞) (ギリシャ語から。 マクロ- 大きい、長い、 ファゴス- むさぼり食う)は、体の防御システムの不均一に特化した細胞集団です。 マクロファージには、遊離マクロファージと固定マクロファージの 2 つのグループがあります。 遊離マクロファージには、疎性結合組織または組織球のマクロファージが含まれます。 漿液性腔のマクロファージ。 炎症性浸出液のマクロファージ。 肺の肺胞マクロファージ。 マクロファージは体中を移動することができます。 固定(常駐)マクロファージのグループは、骨髄、骨および軟骨組織(破骨細胞、軟骨細胞)のマクロファージで構成されます。

米。 8.4.マクロファージ:
あ- ラットの皮下結合組織のマクロファージ(顕微鏡写真、鉄ヘマトキシリンで染色)。 b- マクロファージ (電子顕微鏡写真; A.I. Radostina による調製、倍率 18,000): 1 - 核。 2 - 一次リソソーム。 3 - 二次リソソーム。 4 - 小胞体尿細管のプロファイル。 5 - 細胞質の周縁層の微小成長
ゼンク、リンパ節(樹状マクロファージ)、表皮内マクロファージ(ランゲルハンス細胞)、胎盤絨毛マクロファージ(ホフバウアー細胞)、中枢神経系マクロファージ(ミクログリア)。
マクロファージのサイズと形状は、その機能状態に応じて異なります (図 8.4)。 一部を除いて通常はマクロファージ
種(異物の巨大細胞、軟骨細胞および破骨細胞)には 1 つの核があります。 マクロファージの核は、小さく、丸く、豆の形をしており、または不規則な形をしています。 それらには大きなクロマチンの塊が含まれています。 細胞質は好塩基性で、リソソーム、ファゴソーム (特徴)、および飲作用小胞が豊富で、適度な量のミトコンドリア、顆粒小胞体、ゴルジ複合体、グリコーゲン、脂質などの含有物を含んでいます (図 8.4、b を参照)。 マクロファージの細胞質では、「細胞周縁部」が区別され、これによりマクロファージは移動し、細胞質の微小突起を引き込み、エンドサイトーシスおよびエキソサイトーシスを実行する能力が得られます。 プラズマレンマの直下には、直径 5 ~ 6 nm のアクチン フィラメントのネットワークがあります。 直径 20 nm の微小管がこのネットワークを通過し、形質膜に付着します。 微小管は細胞中心から細胞周辺に向かって放射状に伸びており、リソソーム、微小飲作用小胞、その他の構造の細胞内移動に重要な役割を果たしています。 原形質膜の表面には、腫瘍細胞、赤血球、T リンパ球および B リンパ球、抗原、免疫グロブリン、ホルモンの受容体があります。 免疫グロブリン受容体の存在は、免疫反応へのマクロファージの関与を決定します(第 14 章を参照)。
マクロファージの保護機能の発現形態: 1) 異物の吸収とさらなる分解または分離。 2) 直接接触すると中和されます。 3) 異物に関する情報を、それを中和できる免疫担当細胞に伝達する。 4) 身体の防御システムの別の細胞集団に刺激効果を与える。 マクロファージは、異物、抗菌物質、その他の生物学的活性物質(プロテアーゼ、酸加水分解酵素、パイロジェン、インターフェロン、リゾチームなど)を細胞内および細胞外で分解するための酵素を合成する細胞小器官を備えています。
マクロファージの数とその活動は、特に次のように増加します。 炎症過程。 マクロファージは白血球の走化性因子を生成します。 マクロファージによって分泌される IL-1 は、内皮への白血球の接着、好中球によるリソソーム酵素の分泌とその細胞毒性を増加させることができ、リンパ球の DNA 合成を活性化します。 マクロファージは、B リンパ球による免疫グロブリンの産生と T リンパ球と B リンパ球の分化を活性化する因子を生成します。 細胞溶解性抗腫瘍因子、および自身の集団の細胞の再生と分化に影響を与える成長因子は、線維芽細胞の機能を刺激します(第 14 章を参照)。
マクロファージが抗原と接触すると、グルコース消費、脂質代謝、および食作用活性が急激に増加します。
マクロファージは、HSC だけでなく、前単球や単球からも形成されます (図 7.15 を参照)。 実験動物における疎性線維性結合組織のマクロファージの完全な再生は、線維芽細胞よりも約 10 倍速く起こります。
マクロファージの種類の 1 つは、以前は「外来巨細胞」と呼ばれていた多核巨細胞です。
特に異物の存在下で形成される可能性があるため、「身体」。 多核巨細胞は 10 ~ 20 個以上の核を含み、単核マクロファージの融合によって、または細胞切開を伴わない子宮内膜分裂によって発生します。 電子顕微鏡によると、多核巨細胞には合成および分泌装置と多数のリソソームが含まれています。 プラズマレンマは多数の襞を形成します。
マクロファージ システムの概念。この系には、疎性線維性結合組織のマクロファージ (組織球)、肝臓の類洞血管の星状細胞、造血器官 (骨髄、脾臓、リンパ節) の遊離および固定マクロファージ、肺のマクロファージ、炎症性浸出液 (腹膜) が含まれます。マクロファージ)、破骨細胞、異物の巨細胞、神経組織の体およびグリアマクロファージ(ミクログリア)。 それらはすべて活性のある食細胞です。 貪食された物質は細胞内で酵素切断を受け(「完全貪食」)、局所的に発生したり外部から侵入した身体に有害な物質が除去されます。 この細胞は表面に免疫グロブリン受容体を持ち、骨髄前単球と血液単球から生じます。 このような「専門的な」食細胞とは対照的に、線維芽細胞、網様細胞、内皮細胞、好中球顆粒球などは通性吸収能力を持っていますが、これらの細胞はマクロファージ系の一部ではありません。
I. I. メチニコフは、進化の過程で細胞内消化の一形態として生じ、多くの細胞に割り当てられている食作用が、同時に重要な保護機構であるという考えに最初に到達した。 彼はそれらを 1 つのシステムに組み合わせる実現可能性を実証し、それをマクロファージと呼ぶことを提案しました。 マクロファージ システムは、身体の一般的および局所的な保護反応の両方に関与する強力な保護装置です。 生物全体において、マクロファージ系は局所機構と神経系および内分泌系の両方によって調節されています。
1930 ~ 1940 年代。 この保護システムは細網内皮と呼ばれていました。 で 最近これは単核食細胞系と呼ばれますが、この系に含まれる細胞の中には多核食細胞(破骨細胞)も含まれるため、正確に特徴づけることはできません。
肥満細胞(マスト細胞、組織好塩基球、マスト細胞)。 これらの用語は、好塩基性顆粒球の顆粒を思わせる特定の粒度を持つ細胞質内の細胞を指します。 マスト細胞は、炎症、免疫反応の過程で、血液凝固の減少、血液組織関門の透過性の増加に関与します。
ヒトでは、緩い結合組織の層があるところならどこでも、マスト細胞が見られます。 肥満細胞は、消化管の壁、子宮、乳腺、胸腺、扁桃腺に特に多く存在します。 それらは多くの場合、毛細血管、細動脈、細静脈、小さなリンパ管などの微小血管系の血管に沿ってグループで位置しています(図8.5、図8.5)。 A)。

米。 8.5。肥満細胞: A -皮下結合組織内(顕微鏡写真): 1 - 核。 2 - 細胞質内の異染性顆粒。 b- 超微細構造の図(Yu. I. Afanasyevによる): 1 - コア。 2 - ゴルジ複合体。 3 - リソソーム。 4 - ミトコンドリア。 5 - 小胞体。 6 - 微絨毛。 7 - 不均一な顆粒。 8 - 細胞間物質の分泌顆粒
マスト細胞の形状はさまざまです。 細胞の形状は不規則で、楕円形である場合があります。 これらの細胞には、アメーバ状の運動能力があるため、短くて幅広い突起がある場合があります。 人間の場合、このような細胞の幅は4〜14ミクロンの範囲であり、長さは最大22ミクロンです。 細胞核は比較的小さく、通常は円形または楕円形で、クロマチンが密に詰まっています。 細胞質には多数の顆粒があります。 顆粒のサイズ、組成、量は異なります。 それらの直径は約0.3〜1ミクロンです(図8.5、b)。 少数の顆粒はアズール親和性リソソームを正色染色しています。 ほとんどの顆粒は異色症を特徴とし、ヘパリン、コンドロイチン硫酸タイプ A および C、ヒアルロン酸、ヒスタミン、セロトニンを含んでいます。 顆粒は網状、層状、晶質および混合構造を持っています。
マスト細胞の細胞小器官(ミトコンドリア、ゴルジ複合体、小胞体)はあまり発達していません。 細胞質には、プロテアーゼ、リパーゼ、酸性ホスファターゼ、アルカリ性ホスファターゼ、ペルオキシダーゼ、シトクロムオキシダーゼ、ATPアーゼなどのさまざまな酵素が存在します。ただし、ヒスチジンからヒスタミンを合成するために使用されるヒスチジンデカルボキシラーゼは、マーカー酵素として考慮する必要があります。
マスト細胞は顆粒を分泌および放出することができます。 マスト細胞の脱顆粒は、生理学的状態の変化や病原体の作用に反応して発生する可能性があります。 しかし、マスト細胞からの生体アミンの放出は、可溶性成分の分泌(ヒスタミンの分泌)によっても起こる可能性があります。 ヒスタミンは直ちに毛細血管の拡張を引き起こし、その透過性を高め、局所的な浮腫として現れます。 また、降圧作用もあり、炎症の重要なメディエーターでもあります。
ヒスタミンは、細胞間物質の透過性と血液凝固を低下させるヘパリンの拮抗薬として作用し、抗炎症作用があります。
マスト細胞の数は体の生理学的状態に応じて変化します。マスト細胞は、妊娠中は子宮と乳腺で増加し、消化が活発になると胃、腸、肝臓で増加します。
マスト細胞前駆体は赤色骨髄造血幹細胞に由来します。 マスト細胞の有糸分裂のプロセスは非常にまれに観察されます。
形質細胞(形質細胞)。これらの細胞は、抗原が体内に出現したときに、抗体であるガンマグロブリン(タンパク質)の産生を確実にします。 それらはリンパ器官内で B リンパ球から形成され (第 14 章を参照)、通常は中空器官の粘膜の固有層の緩い結合組織、大網、さまざまな腺 (乳腺、唾液など) の間質性結合組織に見られます。 )、リンパ節、脾臓、骨髄など。
形質細胞のサイズは 7 ~ 10 ミクロンの範囲です。 細胞の形状は円形または楕円形です。 穀粒は比較的小さく、円形または楕円形で、偏心して位置しています。 細胞質は非常に好塩基性で、よく発達した同心円状に位置する顆粒小胞体を含んでいます。

米。 8.6.脂肪細胞:
1 - 毛細管; 2 - 脂質封入体。 3 - コア。 4 - 細胞質(顕微鏡写真)。 染色 - 鉄ヘマトキシリン
タンパク質(抗体)が合成されるチックネットワーク。 好塩基球菌は、核に近い細胞質の小さな明るい領域にのみ存在し、いわゆる球体または中庭を形成します。 中心小体とゴルジ複合体はここにあります。 免疫グロブリンの蓄積が好酸性物質の形で見られることがあります。 ルーセル小体。
形質細胞は、抗体の合成と分泌が高いという特徴があり、これが以前の細胞と区別されます。 よく発達した分泌装置により、毎秒数千個の免疫グロブリン分子の合成と分泌が可能になります。 形質細胞の数は、さまざまな感染症、アレルギー性疾患、炎症性疾患で増加します。
脂肪細胞(脂肪細胞)。これは、栄養、エネルギー生産、水分代謝に関与する、予備脂肪を大量に蓄積する能力のある細胞の名前です。 脂肪細胞はグループで存在し、単独で存在することは少なく、通常は血管の近くに存在します。 これらの細胞は大量に蓄積すると、脂肪組織(白色または茶色)を形成します(下記を参照)。
単一脂肪細胞の形状は球形です。 成熟した脂肪細胞には通常、大きな一滴の中性脂肪 (トリグリセリド) が含まれており、細胞の中央部分全体を占め、薄い細胞質の縁に囲まれており、その厚くなった部分に核があります (図 8.6)。 さらに、脂肪細胞の細胞質には、コレステロール、リン脂質、遊離脂肪酸などの他の脂質も少量含まれています。脂質は、オレンジ色のスダン III または黒色のオスミン酸でよく染色されます。 核に隣接する細胞質内、および場合によってはその反対側の薄い部分にある

米。 8.7.皮膚の色素細胞(顕微鏡写真)。 製剤は着色されていません
高密度に位置するクリステを有する棒状および糸状のミトコンドリアが明らかになります。 多数の飲作用小胞が細胞周囲に見られます。
褐色脂肪組織の脂肪細胞は、核の周囲に位置する小さな脂質封入体とミトコンドリアによって区別されます。 脂肪細胞は優れた代謝能力を持っています。 脂肪細胞内の脂肪封入体の数と疎性結合組織内の脂肪細胞自体の数は両方とも、大幅に変動します。
脂肪細胞の機能は栄養的であり、体内のエネルギーと水分の貯蔵、および体温調節のプロセスへの参加に関連しています。
成人の体の結合組織内の新しい脂肪細胞は、毛細血管に隣接する外膜 (形成層) 細胞からの栄養が増加すると発達します。 この場合、脂肪の小さな液滴が最初に細胞の細胞質に現れ、サイズが大きくなり、徐々に大きな液滴と合体します。 脂肪の減少が増加すると、小胞体とゴルジ複合体が減少し、核が圧縮され、平らになり、細胞の周囲に押し出されます。
多くの臓器に見られる脂肪細胞の塊は脂肪組織と呼ばれます。
外膜細胞。これらは、小さな血管に伴う低分化(形成層)細胞です。 それらは、わずかに好塩基性の細胞質、楕円形の核、および少数の細胞小器官を備えた、平らまたは紡錘形の形状をしています。 これらの細胞は、分岐分化を通じてさまざまな細胞分化(線維芽細胞、筋線維芽細胞、脂肪細胞など)を生じます。
周皮細胞- 毛細血管を取り囲み、その壁を構成する細胞(第 13 章を参照)。
色素細胞(色素細胞、メラノサイト)。 これらの細胞は細胞質内にメラニン色素を含んでいます。 それらの多くは あざ、黒人と黄色人種の人々の結合組織にも存在します。 色素細胞は、短く不規則な形状の突起 (図 8.7)、15 ~ 25 nm の多数のメラノソーム (メラニン顆粒) およびリボソームを持っています。 メラノサイトからの一部のメラノソームは、表皮の有棘層および基底層のケラチノサイトに移動します。
メラノサイトの細胞質には生物学的に活性なアミンも含まれており、マスト細胞とともに血管壁の緊張の調節に関与します(第 14 章を参照)。
メラノサイトは、結合組織内に位置するため、形式的には結合組織にのみ属します。 それらの起源に関しては、これらの細胞は間葉からではなく、神経堤の幹細胞から形成されることが証明されています。 結合組織細胞は、特に炎症や外傷後の再生の過程において、体の水-塩分体制などに違反して、多数の相互作用因子により機能的に単一のシステムに接続されています。
細胞間物質
細胞間物質とか、 マトリックス (細胞間質)、結合組織は、コラーゲンと弾性線維、および粉砕(非晶質)物質から構成されます。 胚と成体の両方の細胞間物質は、一方では結合組織細胞によって行われる分泌によって、他方では細胞間隙に入る血漿から形成されます。
ヒトの胎児では、子宮内発育の 1 ~ 2 か月目から細胞間物質の形成が始まります。 生きている間、細胞間物質は常に新しく生まれ変わります。
人間や動物の結合組織を構成するコラーゲン構造は、その最も代表的な構成要素であり、複雑な組織階層を形成しています。 コラーゲン構造のグループ全体の基礎は繊維状タンパク質です。 コラーゲン、それはコラーゲン構造の特性を決定します。
コラーゲン線維(コラーゲン線維)さまざまな種類の結合組織の組成がその強度を決定します。 緩い未形成の結合組織では、それらは、厚さ1〜3ミクロン以上の波状に湾曲した、らせん状にねじれた、丸い、または平らなストランドの形でさまざまな方向に位置しています。 長さが違います。 コラーゲン線維の内部構造は、線維芽細胞の顆粒小胞体のリボソーム上で合成される線維状タンパク質であるコラーゲンによって決定されます。
コラーゲンには約 20 種類あり、分子構成、器官および組織の所属が異なります。

米。 8.8。コラーゲン生合成と線維形成
人体では、最も一般的なものは次のとおりです。I 型コラーゲンは主に皮膚、腱、骨、角膜、強膜、動脈壁の結合組織に存在します。 II 型コラーゲンは硝子軟骨、線維軟骨、硝子体、角膜の一部です。 III 型コラーゲンは、胎児の皮膚の真皮、大きな血管の壁、造血器官の網状線維に存在します。 IV型 - 基底膜、水晶体嚢内。 V 型コラーゲンは、絨毛膜、羊膜、筋内膜、筋周膜、皮膚、コラーゲンを合成する細胞 (線維芽細胞、内皮、平滑筋) の周囲に存在します。
コラーゲン分子は長さ約 280 nm、幅 1.4 nm です。 それらは、細胞内で単一のらせん状にねじれた、コラーゲン前駆体であるプロコラーゲンの 3 つのポリペプチド鎖 (α 鎖) であるトリプレットから構築されています。 これが最初のものです 分子、コラーゲン線維組織のレベル。 プロコラーゲンは細胞間物質に分泌されます(図8.8)。
2番、 超分子、レベル - コラーゲン線維の細胞外組織 - トロポコラーゲン分子は長さ方向に凝集し、水素結合を介して架橋され、末端プロコラーゲンペプチドの切断によって形成されます。 まずプロトフィブリルが形成され、5~6本のプロトフィブリルが側方結合で結合して厚さ約5nmのミクロフィブリルを構成します。
同じく線維芽細胞によって分泌されるグリコサミノグリカンの関与により、3番目が形成されます。 原繊維、コラーゲン線維組織のレベル。 コラーゲン線維は横縞になっています

米。 8.9.コラーゲンフィブリル (N. P. Omelyanenko による調製)。 P - 期間。 電子顕微鏡写真。 20万件増加
平均厚さ 20 ~ 100 nm の構造。 暗い部分と明るい部分の繰り返し周期は 64 ~ 67 nm です (図 8.9、8.10)。 平行な列の各コラーゲン分子は、隣接する鎖に対して長さの 4 分の 1 だけオフセットされており、これが交互に現れる暗い縞と明るい縞を引き起こしていると考えられています。 電子顕微鏡下の暗いバンドでは、コラーゲン分子内の極性アミノ酸の配置によって引き起こされる二次的な細い横線が見えます。
第4、 ファイバ、組織のレベル。 原線維の集合によって形成されるコラーゲン線維は、1 ~ 10 μm の厚さ(地形に応じて異なります)を持っています。 これには、単一から数十までのさまざまな数のフィブリルが含まれています。 繊維は最大 150 ミクロンの厚さの束に折りたたむことができます。
コラーゲン繊維は、伸びが低く、引張強度が高いという特徴があります。 水中では、膨潤の結果として腱の厚さは50%増加し、希酸および希アルカリ中では10倍増加しますが、繊維は30%短くなります。 膨張する能力は、若い繊維でより顕著になります。 水中で熱処理すると、コラーゲン線維が接着物質を形成します(ギリシャ語: コラ- 接着剤)、これらの繊維に名前が付けられました。
コラーゲン線維の種類は、 網状のそして プレコールラゲン繊維。後者は、胚発生および再生中のコラーゲン線維形成の初期形態を表します。 これらには、III 型コラーゲンと大量の炭水化物が含まれており、造血器官の網様細胞によって合成されます。 それらは三次元ネットワーク、つまり網状組織を形成し、これが名前を決定します。
弾性繊維。弾性繊維の存在 (弾性線維)結合組織の弾力性と伸長性が決まります。 弾性線維はコラーゲン線維に比べて強度が劣ります。 繊維の断面形状は丸くて扁平です。 疎性結合組織では、それらは互いに広く吻合します。 弾性線維の厚さは通常、コラーゲンよりも薄い (0.2 ~ 1 ミクロン) が、(例えば項部靱帯などでは) 数ミクロンに達することもあります。 弾性繊維の組成は次のように分けられます。 ミクロフィブリルそして まとまりのないコンポーネント。

米。 8.10。細胞間物質の形成 (R. Krstic による、修正あり):
1 - 線維芽細胞。 2 - ポリペプチド鎖。 3 - トロポコラーゲン分子; 4 - グリコサミノグリカン。 5 - トロポコラーゲン分子の重合。 6 - プロトフィブリル。 7 - プロトフィブリルの束(コラーゲンフィブリル)。 8 - エラスチン分子。 9 - 弾性フィブリル
弾性線維の基礎は球状糖タンパク質です - エラスチン、線維芽細胞と平滑筋細胞によって合成されます(まず、分子、組織レベル)。 エラスチンは、高含有量のプロリンとグリシン、およびエラスチンの分子構造を安定化し、伸縮性と弾力性を与える 2 つのアミノ酸誘導体、デスモシンとイソデスモシンの存在によって特徴付けられます。 細胞の外側にある直径 2.8 nm の小球を持つエラスチン分子は鎖状につながっており、厚さ 3 ~ 3.5 nm のエラスチン プロトフィブリル (2 番目、超分子、組織レベル) です。 エラスチンプロトフィブリルは、糖タンパク質(フィブリリン)と結合して、厚さ8〜19 nmのミクロフィブリル(図8.11)を形成します(3番目、フィブリル、組織レベル)。 組織の第 4 レベルは、

米。 8.11。弾性繊維の超微細構造: あ- 図: 1 - 中央の均質部分。 2 - ミクロフィブリル(Yu. I. Afanasyevによる)。 b- 電子顕微鏡写真、倍率 45,000 (V.P. Slyusarchuk による作成): 1 - 中央の均質部分。 2 - 繊維の周囲にあるミクロフィブリル。 3 - 線維芽細胞ゴルジ複合体。 4 - 小胞体。 5 - 中心小体
ファイバ。 最も成熟した弾性繊維は、中心に約 90% の非晶質弾性タンパク質成分 (エラスチン) を含み、周囲に沿ってミクロフィブリルを含みます。 弾性線維には、コラーゲン線維とは異なり、その長さに沿って横縞のある構造がありません。
成熟した弾性線維に加えて、 エラウニンそして オキシタラン繊維。 エラウニン繊維では、ミクロフィブリルと非晶質成分の比率がほぼ等しいのに対し、オキシタラン繊維はミクロフィブリルのみで構成されています。
細胞間物質の非晶質成分。結合組織の細胞と繊維は、非晶質成分、つまり基質の中に閉じ込められています。 (実質的原理)。このゲル状物質は、結合組織、神経および血管の細胞構造および線維構造を取り囲む、代謝的で統合的な緩衝作用を有する多成分環境です。 主物質の成分には、血漿タンパク質、水、無機イオン、細胞代謝産物、コラーゲンとエラスチンの可溶性前駆体、プロテオグリカン、糖タンパク質、およびそれらによって形成される複合体が含まれます。 これらすべての物質は絶えず移動し、更新されています(図8.12)。
プロテオグリカン (PG) は、90 ~ 95% の炭水化物を含むタンパク質と炭水化物の化合物です。
グリコサミノグリカン (GAG) は、通常、ヘキスロン酸とアミノ糖 (N-アセチルグリコサミン、N-アセチル銀河酸) を含む多糖化合物です。
プロテオグリカン

米。 8.12非晶質結合組織物質の分子構成
トサミン)。 GAG 分子には、負の電荷を持つヒドロキシル、カルボキシル、硫酸基が多数含まれており、水分子やイオン、特に Na + と容易に結合するため、組織の親水性が決まります。 GAG は酸素と CO 2 を透過しますが、異物やタンパク質の侵入から臓器を保護します。 GAG は、結合組織の繊維構造の形成とその機械的特性、結合組織の修復プロセス、および細胞の成長と分化の制御に関与しています。 これらの化合物の中で、結合組織の種類で最も一般的なのはヒアルロン酸と硫酸化 GAG です。コンドロイチン硫酸 (軟骨、皮膚、角膜)、デルマタン硫酸 (皮膚、腱、血管壁など)。 )、ケラタン硫酸、ヘパリン硫酸(多くの基底膜の組成に含まれる)。
ヘパリンは、グルクロン酸とグリコサミンからなるGAGです。 人間や動物では、肥満細胞によって生成され、血液中の天然の抗凝固因子です。
糖タンパク質 (GP) は、繊維と非晶質物質の両方の一部であるオリゴ糖 (ヘキソース、ヘキソース、フコース、シアル酸) を含むタンパク質化合物のクラスです。 これらには、プロテオグリカンに関連する可溶性 GP が含まれます。 石灰化組織の GP; コラーゲン関連の GP。 GP は結合組織の細胞間物質の構造の形成に主要な役割を果たし、その機能的特徴 (フィブロネクチン、コンドロネクチン、フィブリリン、ラミニンなど) を決定します。
フィブロネクチンは線維芽細胞の主表面 GP です。 細胞間空間では、主に間質コラーゲンと関係しています。 フィブロネクチンは、細胞の粘着性、運動性、成長および特殊化などを決定すると考えられています。
フィブリリンはミクロフィブリルを形成し、細胞外成分間の結合を強化します。
ラミニンは基底膜の構成要素であり、ジスルフィド化合物、V 型コラーゲンおよび細胞表面受容体によって相互接続された 3 つのポリペプチド鎖から構成されます。
密な結合組織
密な結合組織 (テクストゥス・コネクティブス・コンパクトス)比較的多数の高密度に配置された繊維と少量の細胞要素、およびそれらの間の主な非晶質物質によって特徴付けられます。 繊維構造の位置の性質に応じて、それらは区別されます。 方向性のない(未形成)、 指向性のある(フォーマット済み)および 混合された密な結合組織の一種。
方向性のない繊維の無秩序な配置を特徴とする組織の一種 (たとえば、皮膚の真皮)。 で 指向性のある組織の種類に応じて、線維の配置は厳密に順序付けられており、それぞれの場合において臓器(腱、靱帯、線維膜)が機能する状態に対応しています。 混合タイプは、原則として、高密度の結合組織の線維要素が交互の方向にある層状構造(角膜、強膜)を持っています。
腱(天童)。それは、平行に配置されたコラーゲン線維の厚く密に詰まった平行な束で構成されています。 これらの束の間には、と呼ばれる線維細胞が存在します。 腱細胞 (腱細胞)、少量の線維芽細胞と粉砕された非晶質物質。 線維細胞の薄い層状突起が線維束間の空間に入り、線維束と密接に接触します。 隣接する線維細胞の層から分離されたコラーゲン線維の各束は、と呼ばれます。 一次ビーム。緩い結合組織の薄い層に囲まれたいくつかの 1 次の束が 2 次の束を構成します。 分離する疎性結合組織の層 二次ビーム、呼ばれた エンドテノニウム。二次ビームから構成されます 3次ビーム、疎性結合組織の厚い層によって分離されています - ペリテノニウム。場合によっては、三次束が腱自体である場合もあります。 大きな腱には 4 次の束がある場合もあります。
ペリテノニウムとエンドテノニウムには、腱、神経、および腱組織の緊張状態について中枢神経系に信号を送る固有受容神経終末に栄養を供給する血管が含まれています。
項部靱帯も形成されたタイプの密な結合組織に属しますが、それのみが厚い弾性繊維から形成されます。
骨に付着している腱の一部は、2 つの線維性結合組織鞘でできた鞘に包まれています。 (膣)その間にはヒアルロン酸が豊富な液体(潤滑剤)があります。
繊維膜。これらの構造は高密度の結合組織で構築されており、筋膜、腱膜、横隔膜の腱中心、一部の臓器の被膜、硬膜、強膜、軟骨膜、骨膜、さらには卵巣や精巣の白膜などが含まれます。このタイプの組織の線維性膜は、コラーゲン線維の束と、それらの間にある線維芽細胞および線維細胞が、互いにいくつかの層の上に一定の順序で配置されているため、伸ばすことが困難です。 各層では、コラーゲン線維の波形の束が、隣接する層の方向とは一致しない一方向に互いに平行に走っています(異なる方向に平行に組織された結合組織)。 繊維の個々の束が 1 つの層から別の層に移動し、それらを結合します。 コラーゲン線維の束に加えて、線維膜には弾性線維が含まれています。 骨膜、強膜、精巣白膜、関節包などの線維構造は、腱膜と比較して、コラーゲン線維束の規則的でない配置と多数の弾性線維によって特徴付けられます。
8.1.2. 特別な性質を持つ結合組織
このような生地としては、 網状、脂肪質そして 粘膜それらは均質な細胞が優勢であることを特徴としており、これらのタイプの結合組織の名前自体が通常それに関連付けられています。
網状組織
網状組織 (網状組織)結合組織の一種で、ネットワーク状の構造を持ち、プロセスで構成されています。 網様細胞そして 網状(好銀性)線維(図8.13)。 ほとんどの網様細胞は網様線維と結合しており、突起によって互いに接続され、三次元ネットワークを形成しています。 網状組織は、造血器官の間質と、そこで発達する血球の微環境を形成します。
網状線維 (直径 0.5 ~ 2 μm) は、網状細胞の合成の産物です。 銀塩を含浸させると検出されるため、親銀性(ギリシャ語から)とも呼ばれます。 アルゲンタム- シルバー)。 これらの繊維は弱酸や弱アルカリに耐性があり、トリプシンによって消化されません。
好銀性線維のグループでは、網状線維とプレコラーゲン線維が区別されます。 網状線維自体は、III 型コラーゲンを含む最終的な最終形成物です。 網状線維は、コラーゲン線維と比較して、高濃度の硫黄、脂質、炭水化物を含んでいます。 電子顕微鏡では、網状線維フィブリルには、周期 64 ~ 67 nm の明確に定義された縞模様があるとは限りません。 伸長性の観点から、これらの繊維は
コラーゲンとエラスティックの中間の位置にあります。 プレコラーゲン線維は、胚形成および再生中のコラーゲン線維形成の初期形態を表します。

米。 8.13。網状組織: あ- 網様細胞の顕微鏡写真: 1 - 網様細胞の核。 2 - 細胞質のプロセス。 b- リンパ節の網状線維の顕微鏡写真: 1 - 網状線維。 硝酸銀の含浸
脂肪組織
脂肪組織 (脂肪組織)- これらは多くの臓器に見られる脂肪細胞の蓄積です (脂肪細胞を参照)。 脂肪組織には 2 種類あります - 白そして 茶色。これらの用語は条件付きであり、セルの色の特性を反映しています。 白色脂肪組織は人体に広く分布しており、褐色脂肪組織は主に新生児と一部の動物(げっ歯類や冬眠動物)に生涯を通じて見られます。
白色脂肪組織ヒトでは、皮下、特に腹壁の下部、臀部および大腿部に存在し、そこで皮下脂肪層を形成し、大網、腸間膜および後腹膜領域に存在します。 脂肪組織は、疎性線維性結合組織の層によって、さまざまなサイズと形状の小葉に多かれ少なかれ明確に分割されています。 小葉内の脂肪細胞は互いに非常に近接しています (図 8.14)。 線維芽細胞、リンパ系要素、および肥満細胞は、それらの間の狭い空間に位置しています。 脂肪細胞の間には細いコラーゲン線維が全方向に配向しています。 疎性線維性結合組織の層に位置する血液およびリンパの毛細管は、脂肪細胞のグループまたは脂肪組織の小葉をそのループで密に取り囲んでいます。 脂肪組織では、脂肪酸、炭水化物の代謝、および炭水化物からの脂肪の形成の活発なプロセスが発生します。
脂肪の分解中に、大量の水が放出され、エネルギーが放出されるため、脂肪組織は、高エネルギー化合物の合成のための基質の貯蔵庫としての役割だけでなく、間接的に水の貯蔵庫としての役割も果たします。 。
絶食中、皮下および腎周囲脂肪組織、大網および腸間膜脂肪組織は、蓄積した脂肪を急速に失います。 脂質滴
細胞内の脂肪が粉砕され、脂肪細胞は星状または紡錘状の形状になります。 目の眼窩領域や手のひらや足の裏の皮膚では、長期間の絶食中でも脂肪組織は少量の脂質しか失いません。 ここで、脂肪組織は代謝というよりも主に機械的な役割を果たします。 これらの場所では、結合組織線維に囲まれた小さな小葉に分割されます。
褐色脂肪組織新生児や一部の動物の首、肩甲骨の近く、胸骨の後ろ、脊椎に沿って、皮膚の下、筋肉の間に見られます。 それは毛細血管と密に絡み合った脂肪細胞で構成されています。 これらのセルは熱生成プロセスに関与します。 褐色脂肪組織の脂肪細胞は、細胞質に多くの小さな脂肪封入体を持っています(図8.15)。 白色脂肪組織細胞と比較して、有意に多くのミトコンドリアを持っています。 脂肪細胞の茶色は、鉄を含む色素、ミトコンドリアのシトクロムによって与えられます。
褐色脂肪細胞の酸化能力は白色脂肪細胞の約 20 倍、心筋の酸化能力のほぼ 2 倍です。 周囲温度が低下すると、褐色脂肪組織の酸化プロセスの活動が増加します。 で

米。 8.14。白色脂肪組織の構造 (Yu. I. Afanasyev による): あ b- 脂肪細胞の超顕微鏡構造: 1 - 脂肪細胞核; 2 - 脂質の大滴。 3 - 神経線維。 4 - 毛細血管。 5 - 脂肪細胞ミトコンドリア。 V- 脂肪組織: 1 - 脂肪細胞; 2 - 血管。 顕微鏡写真(スーダンIII染色)

米。 8.15。褐色脂肪組織の構造 (Yu. I. Afanasyev による): あ- 光学光学顕微鏡で観察した脂肪を除去した脂肪細胞。 b- 脂肪細胞の超顕微鏡構造: 1 - 脂肪細胞核; 2 - 細かく砕いた脂質。 3 - 多数のミトコンドリア。 4 - 毛細血管。 5 - 神経線維
これにより熱エネルギーが放出され、毛細血管内の血液が温められます。 熱交換の調節では、交感神経系と副腎髄質のホルモンであるアドレナリンとノルアドレナリンが一定の役割を果たします。これらは、トリグリセリドをグリセロールに分解する組織リパーゼの活性を刺激します。 脂肪酸。 これにより熱エネルギーが放出され、脂肪細胞間の多数の毛細血管を流れる血液が温められます。 絶食中、褐色脂肪組織は白色脂肪組織よりも変化が少なくなります。
粘膜組織
粘膜組織(粘膜組織)通常は胚の中でのみ発生します。 研究の古典的な対象は人間の胎児のへその緒です (図 8.16)。 ここでの細胞要素は、出生前期間中に間葉細胞から分岐して分化した不均一な細胞群によって表されます。 その中で、大きな星型線維芽細胞、筋線維芽細胞、平滑筋細胞が区別されます。 それらは、ビメンチン、デスミン、アクチン、およびミオシンを合成する能力によって区別されます。 臍管の粘膜組織の線維芽細胞 -

米。 8.16。臍帯からの粘液結合組織: 1 - 粘液細胞。 2 - 細胞間物質。 3 - 血管壁
チーク (「ウォートンのゼリー」) は、基底膜の特徴である IV 型コラーゲン、ラミニン、ヘパリン硫酸を合成します。
妊娠の前半には、この組織の細胞間にヒアルロン酸が大量に存在し、主物質がゼリー状になります。 ゼラチン状結合組織の線維芽細胞は、線維状タンパク質を弱く合成します。 胚発生の後期段階になって初めて、緩く配置されたコラーゲン原線維がゼラチン状物質の中に現れます。
8.2. 骨格組織
骨格組織 (テクストゥス スケルトン)- これらは、高密度の細胞間物質(軟骨、骨組織、象牙質、歯科用セメント)の存在により、顕著な支持力、機械的機能を持つ結合組織の一種です。 これらの組織は、主な機能に加えて、水と塩の代謝にも関与します。
8.2.1. 軟骨組織
軟骨組織 (軟骨組織)呼吸器系、関節、椎間板などの器官の一部であり、細胞で構成されています。 軟骨細胞そして 軟骨芽細胞細胞間に親水性物質が多く含まれており、弾力性が特徴です。 これには軟骨組織の支持機能が関係しています。 新鮮な軟骨組織には、約 70 ~ 80% の水分、10 ~ 15% の有機物質、および 4 ~ 7% の塩が含まれています。 軟骨組織の乾燥物質の 50 ~ 70% はコラーゲンです。 軟骨組織自体には血管がなく、栄養は周囲の組織から拡散します。 軟骨膜。
分類。軟骨組織には 3 つのタイプがあります。 ヒアリン、弾性、繊維質。軟骨組織のこの分割は、細胞間物質の構造、コラーゲンと弾性線維の含有量と比率の構造的および機能的特徴に基づいています。
軟骨組織の発生(軟骨形成)
軟骨組織の発生源は間葉です。 最初に、軟骨が形成される胚の体の一部の部分では、間葉細胞が
私たちはプロセスを失い、活発に増殖し、互いにしっかりと隣接して、ある種の緊張、つまり膨満感を生み出します。 このような領域はこう呼ばれます 軟骨形成の基礎、または 軟骨形成島(図8.17)。 それらには、半幹軟骨形成細胞に分化する幹細胞が含まれています。 幹細胞は、丸い形、高い値の核細胞質比、拡散したクロマチンの配置、および小さな核小体によって特徴付けられます。 これらの細胞内の細胞小器官は十分に発達していません。 半幹細胞(前軟骨芽細胞)では、遊離リボソームの数が増加し、顆粒型の小胞体膜が現れ、細胞の形状が長くなり、核と細胞質の関係が減少します。 その後、低分化前駆体は形態学的に識別可能な軟骨芽細胞に分化します (図 8.17 を参照)。
軟骨芽細胞(ギリシャ語から。 コンドロ- 軟骨、 爆風- 原始的)は、増殖し、軟骨の細胞間物質(プロテオグリカン)を合成できる若い平らな細胞です。 軟骨芽細胞の細胞質には、よく発達した顆粒および無顆粒小胞体であるゴルジ複合体があります。 染色すると、軟骨芽細胞の細胞質は豊富な含有量により好塩基性であることがわかります。
米。 8.17。硝子軟骨組織の発達 (Yu. I. Afanasyev によるスキーム): あ- 軟骨形成島; b- 一次軟骨組織; V- 軟骨細胞の分化の段階。 1 - 間葉細胞。 2 - 有糸分裂細胞。 3 - 細胞間物質。 4 - 好酸性細胞間物質。 5 - 好塩基性細胞間物質。 6 - 若い軟骨細胞。 7 - 毛細血管

米。 8.18。軟骨細胞の同質遺伝子群:
1 - 軟骨細胞; 2 - 非コラーゲン性タンパク質とプロテオグリカンのループ状ネットワークからなる細胞領域のマトリックス。 3 - 腔の壁を形成するコラーゲン線維。 4 - 軟骨の領域間部分。 5 - 領土間マトリックス中のプロテオグリカン(V.N. パブロワによる、修正あり)
RNA。 軟骨芽細胞は線維状タンパク質(コラーゲン)を合成および分泌し、好酸性染色される細胞間物質が現れます。 これが一次軟骨組織の形成方法です。 軟骨芽細胞の関与により、 周辺(隣接)成長軟骨。 軟骨組織がさらに分化すると、軟骨芽細胞は軟骨細胞に発達します。
軟骨細胞- 軟骨組織細胞の主な種類。 それらは、分化の程度に応じて、楕円形、円形、または多角形の形状になります。 それらは、細胞間物質の特別な空洞(空隙)に、単独またはグループで位置しています。 共通の空洞内にある細胞のグループは、 同質遺伝子型(ギリシャ語から。 アイソ- 等しい、 創世記- 発達)。 それらは1つの細胞の分裂によって形成されます(図8.18)。 同質遺伝子群では、3 種類の軟骨細胞が区別されます。
最初のタイプの軟骨細胞は、高い核細胞質比、ゴルジ複合体の空胞要素の発達、細胞質内のミトコンドリアと遊離リボソームの存在によって特徴付けられます。 これらの細胞では、有糸分裂の様子がしばしば観察されるため、それらを同質遺伝子型の細胞グループの出現の源と考えることができます(図8.19)。 I 型軟骨細胞は、発達中の若い軟骨で優勢です。 II 型軟骨細胞は、核細胞質比の減少、DNA 合成の弱化、高レベルの RNA の保存、顆粒小胞体とゴルジ複合体のすべての構成要素の集中的な発達によって特徴づけられ、それらは細胞質の形成と分泌を確実にします。グリコサミノグリカンとプロテオグリカンを細胞間物質に取り込みます。 III 型軟骨細胞は、核と細胞質の比率が最も低く、顆粒小胞体の強力な発達と秩序ある配置によって特徴付けられます。 これらの細胞はタンパク質を形成および分泌する能力を保持していますが、グリコサミノグリカンの合成は減少しています。
間葉との境界の周縁部に軟骨原基が発達すると、 軟骨膜- 発達中の軟骨を外側から覆い、外側の繊維層と内側の軟骨形成層(形成層)からなる殻。 軟骨形成領域では、細胞が集中的に分裂し、軟骨芽細胞に分化します。軟骨芽細胞は、DNA を合成する能力を保持し、複製し、細胞間物質 (I 型および III 型コラーゲン) の成分も合成します。 合成産物が分泌され、その周囲に沿って既存の軟骨に層を重ねる過程で、それら自体が
細胞は、その活動の産物で「壁に囲まれ」ています。 このようにして軟骨は重なり合って成長します。 同位的な成長。
若い発達中の軟骨の中心にある軟骨細胞は、しばらくの間有糸分裂する能力を保持し、同じ空隙(細胞の同質遺伝子群)に留まり、II型コラーゲンを産生します。 これらの細胞の数が増加し、細胞間物質が生成されることにより、内側から軟骨の量が増加します。 間質成長。間質の成長は、胚発生時だけでなく、軟骨組織の再生時にも観察されます。
軟骨の成長と発達に伴って、その中央部分は近くの血管からますます遠ざかり、軟骨膜の血管からびまん性に行われる栄養補給が困難になり始めます。 その結果、軟骨細胞は再生能力を失い、その一部は破壊され、プロテオグリカンはより単純な好酸性タンパク質であるアルブロイドに変換されます。
したがって、胚の組織形成における軟骨芽細胞の差異は、あらゆる形態の成熟細胞によって表されます。 ただし、その後は、中間および最後のリンク (軟骨芽細胞および軟骨細胞) のみが含まれ、軟骨組織ではこれが唯一のものになります。

米。 8.19。 3 種類の軟骨細胞 (Yu. I. Afanasyev による):
あ- タイプ I; b -タイプ II; V -Ⅲ型。 1 - 有糸分裂細胞。 2 - 小胞体。 3 - ミトコンドリア。 4 - グリコーゲン。 5 - 細胞間物質
硝子軟骨組織
硝子軟骨組織 (textus cartilagineus hyalinus)、硝子体とも呼ばれます(ギリシャ語から。 ヒアロス- ガラス) - 透明で青みがかった白色のため、最も一般的な品種です -

米。 8.20。硝子軟骨: あ- 気管の硝子軟骨の顕微鏡写真:1 - 軟骨膜。 2 - 若い軟骨細胞。 3 - 内部に同質遺伝子群の軟骨細胞が存在する塩基性物質 (4)
軟骨組織の密度。 成人の身体では、硝子組織は肋骨と胸骨の接合部、喉頭、気道、骨の関節面に見られます。
さまざまな臓器の硝子軟骨組織には多くの共通点がありますが、同時に臓器の特異性、つまり細胞の位置、細胞間物質の構造が異なります。 人体にある硝子軟骨組織のほとんどが覆われています 軟骨膜解剖学的構造を表します - 軟骨。
軟骨膜には 2 つの層があります。外層は血管との線維性結合組織から構成されます。 内部に存在し、主に細胞性であり、軟骨芽細胞とその前駆体であるプレコンドロ芽細胞を含んでいます。 表層の軟骨膜の下には若い紡錘形の軟骨細胞があり、その長軸は軟骨の表面に沿っています(図8.20)。 より深い層では、軟骨細胞は楕円形または円形の形状になります。 これらの細胞の合成プロセスと分泌プロセスが弱まっているという事実により、分裂後、それらは大きく分岐せず、コンパクトに横たわって、2〜4個の軟骨細胞のいわゆる同質遺伝子グループを形成します。
より分化した軟骨細胞および同質遺伝子群は、細胞間物質の好酸性細胞周囲層に加えて、外側に好塩基性ゾーンを持っています。 これらの特性は、細胞間物質の化学成分であるタンパク質とグリコサミノグリカンの不均一な分布によって説明されます。
あらゆる局在の硝子軟骨では区別するのが通例です 領土細胞間物質、またはマトリックス (図 8.18 を参照)。 領域領域には、軟骨細胞またはそのグループのすぐ周囲のマトリックスが含まれます。 これらの領域では、II 型コラーゲン線維と原線維がねじれて同質遺伝子群の軟骨細胞を取り囲み、機械的圧力から細胞を保護しています。 領土内

米。 8.20。継続
b- 軟骨と軟骨膜の構造の図(図。Yu. I. Afanasyev): 1 - 外側の繊維層。 2 - 内部細胞層。 3 - 軟骨組織。 V- 関節軟骨の細胞および繊維成分(V.P. Modyaev、V.N. Pavlovaによる、修正あり)。 I - 表面ゾーン。 II - 中間ゾーン。 III - 基底(深部)ゾーン。 IV - 軟骨下骨。 A - 関節軟骨の細胞成分: 1 - 無細胞プレート。 2 - 接線層の軟骨細胞。 3 - 移行領域の軟骨細胞。 4 - 同質遺伝子グループ。 5 - 軟骨細胞の「柱」。 6 - 肥大した軟骨細胞。 7 - 石灰化軟骨と非石灰化軟骨の間の好塩基性(境界)線。 8 - 軟骨の石灰化。 B - 関節軟骨の線維系: 1 - 無細胞板。 2 - 表面ゾーンの接線方向の繊維。 3 - 中間ゾーンのコラーゲン線維の主な方向。 4 - 基底層の放射状繊維。 5 - 好塩基性(境界)線
このマトリックスでは、コラーゲン線維は主荷重の力の作用ベクトルの方向に配向します。 コラーゲン構造間の空間はプロテオグリカンで満たされています。
コンドロネクチンは、軟骨の細胞間物質の構造組織において重要な役割を果たします。 この糖タンパク質は、細胞同士を接続したり、さまざまな基質 (コラーゲン、グリコサミノグリカン) に接続したりします。 圧縮および張力中の軟骨組織のサポート生体力学的機能は、その線維フレームの構造だけでなく、高レベルの水和(65 ~ 85%)を備えた親水性プロテオグリカンの存在によっても確保されています。 細胞間物質の高い親水性により、栄養素や塩分の拡散が促進されます。 ガスや多くの代謝物もそこを自由に拡散します。 ただし、抗原性を持つ大きなタンパク質分子は通過しません。 これは、クリニックでの軟骨部分の移植(ある人から別の人への移植)が成功したことを説明しています。 軟骨細胞の代謝は主に嫌気性、解糖性です。
ただし、すべての軟骨が同じように構築されているわけではありません。 関節表面の硝子軟骨の構造上の特徴は、関節腔に面する表面に軟骨膜が存在しないことです。 関節軟骨は、表面、中間、基底の 3 つの境界がはっきりしていないゾーンで構成されています (図 8.20、c を参照)。
関節軟骨の表層には、小さくて平らな、特殊化されていない軟骨細胞があり、その構造は線維細胞に似ています。
中間ゾーンでは、細胞はより大きく、丸い形をしており、代謝的に非常に活発です。大きなミトコンドリア、よく発達した顆粒小胞体、および多数の小胞を持つゴルジ複合体を備えています。
深層(基底)ゾーンは好塩基性線によって非石灰化層と石灰化層に分けられます。 後者は、下にある軟骨下骨からの血管によって貫通されます。 関節軟骨の深部の細胞間物質の特徴は、骨格組織の初期石灰化の場所である直径30nmから1μmの膜構造である高密度のマトリックス小胞の内容です(軟骨に加えて、それらは骨組織および象牙前質に見られます)。 膜構造は、軟骨細胞の形質膜(それぞれ、骨組織の骨芽細胞と象牙前質の象牙芽細胞)の一部が膨らみ、続いて細胞表面から出芽し、石灰化ゾーンに局所的に分布することによって形成されます。 それらは完全な細胞崩壊の産物である可能性もあります。 関節軟骨は、深部の血管からは部分的にのみ栄養が供給されますが、主に関節腔の滑液から栄養が供給されます。
弾性軟骨組織
弾性軟骨組織 (弾性軟骨組織)軟骨基部が曲がりやすい器官(耳介、喉頭の角質軟骨および楔形軟骨など)に見られます。 新鮮で固定されていない状態では、弾性軟骨組織は黄色がかった色であり、硝子組織ほど透明ではありません。 弾性軟骨の一般的な構造によれば、それは硝子軟骨に似ています。 外側は軟骨膜で覆われています。 軟骨細胞(若くて特殊化した) 軟骨細胞)莢膜内に単独で存在するか、同質遺伝子群を形成します。 主な違いの 1 つは
弾性軟骨の重要な特徴は、細胞間物質にコラーゲン線維とともに、細胞間物質を全方向に貫通する弾性線維が存在することです(図8.21)。
軟骨膜に隣接する層から、弾性線維は途切れることなく軟骨膜の弾性線維の中を通過します。 弾性軟骨には、硝子軟骨よりも脂質、グリコーゲン、およびコンドロイチン硫酸が少ない。

米。 8.21。弾性のある軟骨組織。 オルセインで染色した顕微鏡写真: 1 - 軟骨細胞の同質遺伝子群。 2 - 弾性繊維
線維性軟骨組織
線維性軟骨組織 (軟骨繊維線維症)椎間板、半可動性の関節、線維性結合組織(腱、靱帯)から硝子軟骨への移行箇所に位置し、限られた動きには強い張力が伴います。 細胞間物質にはコラーゲンの束が平行に伸びており、
徐々に緩み、硝子軟骨に変化します。 軟骨には軟骨細胞が閉じ込められた空洞があります。 後者は単独で存在するか、小さな同質遺伝子群を形成します。 細胞の細胞質は空胞化していることがよくあります。 硝子軟骨が腱に向かって移動すると、線維軟骨はますます腱に似てきます。 軟骨と腱の境界、コラーゲン束の間には、圧縮された軟骨細胞の列があり、境界もなく、密な結合組織に位置する腱細胞に入ります(図8.22)。
年齢は変わります。身体が老化するにつれて、軟骨組織内のプロテオグリカンの濃度とそれに関連する親水性が減少します。 軟骨芽細胞と若い軟骨細胞の再生プロセスが弱まります。 これらの細胞の細胞質では、ゴルジ複合体、顆粒小胞体、ミトコンドリアの体積が減少し、酵素活性が低下します。
異栄養的に変化した細胞および細胞間物質の吸収に関与する 軟骨細胞、形態的には破骨細胞と同一です。 軟骨細胞が死んだ後、隙間の一部は非晶質物質とコラーゲン原線維で埋められます。 細胞間のところどころに

米。 8.22線維性の軟骨組織。 椎間板のセクション: 1 - コラーゲン線維。 2 - 軟骨細胞
この物質では、カルシウム塩の沈着が検出され(「軟骨のチョーキング」)、その結果、軟骨は曇り、硬くなり、脆くなります。 その結果、軟骨の中央領域の栄養障害が生じ、血管が軟骨内に内方成長し、その後、骨が形成される可能性があります。
再生。軟骨組織の生理学的再生は、軟骨膜および軟骨の未分化細胞により、軟骨芽前細胞および軟骨芽細胞の再生および分化を通じて行われる。 ただし、このプロセスは非常に時間がかかります。 関節外局在の軟骨組織の外傷後の再生は、軟骨膜によって行われます(図8.23)。
関節軟骨では、損傷の深さに応じて、同質遺伝子群の細胞のみの増殖(損傷が浅い場合)と、第二の再生源である軟骨下骨組織の形成層細胞(損傷が深い場合)の両方によって再生が起こります。軟骨への損傷)、骨の有機基質を形成します - 類骨。
いずれの場合でも、ジストロフィー(壊死)プロセスは軟骨組織損傷の領域で直接観察され、増殖中の軟骨芽細胞はより遠心的に位置します。 線維性結合組織が創傷内に形成され、その後軟骨に置き換わります。 一般に、再生は受傷後 3 ~ 6 か月で完了します。
軟骨組織の代謝を調節する因子。軟骨組織の代謝の調節は、機械的負荷、神経およびホルモン因子の影響下で発生します。 軟骨組織への周期的な圧力と負荷の弱化は、受容体とエフェクターを備えた軟骨膜の毛細血管、または関節の滑液から、水に溶解した栄養素、代謝産物、ホルモン・液性調節因子の拡散の要因として常に作用しています。 。 さらに、軟骨細胞は、血液中を循環する多くのホルモンに対する細胞受容体を持っています。 つまり、下垂体ホルモンは成長ホルモンです
プロラクチン - 軟骨組織の成長を刺激しますが、その成熟には影響しません。 甲状腺ホルモンであるチロキシンとトリヨードチロニンは、軟骨細胞の細胞分化を促進しますが、軟骨の成長プロセスを阻害します。 甲状腺と副甲状腺のホルモンであるカルシトニンと副甲状腺ホルモンは、軟骨の代謝に同様の影響を及ぼし、成長プロセスの刺激に役立ちますが、成熟の程度はそれほど高くありません。 膵臓の内分泌島のホルモンであるインスリンは、骨格形成間葉細胞の細胞分化を促進し、出生後の個体発生の段階では、成長および分裂促進効果をもたらします。 副腎皮質のホルモンであるグルココルチコイドと女性ホルモンのエストロゲンは、軟骨細胞内のコラーゲンとグリコサミノグリカンの生合成を阻害し、出生後早期にはそれらの高濃度が軟骨組織の老化とその破壊的な変化に寄与します。 男性ホルモンであるテストステロンは、非硫酸化グリコサミノグリカンの生合成を刺激し、軟骨組織の成熟度の低下を引き起こします。 一般に、ホルモンは軟骨細胞の特定の代謝プロセスを調節しますが、ホルモンの作用に応答する軟骨細胞の能力は、体の内分泌状態(ホルモンの正常、欠乏または過剰)とホルモンの両方に依存することに注意する必要があります。軟骨細胞自体の構造的および機能的状態。
8.2.2. 骨組織
骨組織 (テキサス・オッセイ)細胞間の高度な石灰化を伴う特殊な多種類の結合組織です。

米。 8.23。浅い硝子軟骨の外傷後の再生 (1そして
b 1 と深い (a2 b 2) 損傷: あ\ 2 - 関節軟骨の再生: 1 - 滑膜。 2 - 軟骨。 3 - 骨。 4 - 壊死ゾーン。 5 - 拡散ゾーン。 6 - 非石灰化軟骨。 7 - 石灰化した軟骨。 8 - 血管のあるオステオン。 9 - 骨髄。 10 - 肉芽組織。 b - 肋軟骨の再生:1 - 軟骨膜。 2 - 軟骨。 3 - 壊死ゾーン。 4 - 拡散ゾーン。 5 - 肉芽組織(V.N. Pavlovaによる)。 矢印 - 創傷過程における細胞と組織の移動方向
約70%の無機化合物を含む有機物で、主にリン酸カルシウム。 骨組織には 30 種類以上の微量元素 (銅、ストロンチウム、亜鉛、バリウム、マグネシウムなど) が検出され、体内の代謝プロセスで重要な役割を果たしています。
有機物 - マトリックス骨組織 - 主にコラーゲン型のタンパク質と脂質で表されます。 軟骨組織のマトリックスと比較して、それには比較的少量の水分、コンドロイチン硫酸が含まれていますが、カルシウムと複合体を形成し、骨の有機マトリックスに浸透するクエン酸やその他の酸が多く含まれています。 有機成分と無機成分が互いに組み合わされて、機械的特性、つまり伸長や圧縮に耐える能力が決まります。あらゆる種類の結合組織の中で、支持機能、機械機能、保護機能は骨組織で最も顕著です。 内臓にとっては、カルシウム、リンなどの塩類の貯蔵庫でもあります。
高度な石灰化にもかかわらず、骨組織は、その構成物質の絶え間ない更新、絶え間ない破壊と生成、および変化する動作条件による適応を経ます。 骨組織の形態機能的特性は、年齢、筋肉活動、栄養状態に応じて、また内分泌腺の活動、神経支配などの影響を受けて変化します。
分類。骨組織には主に 2 つのタイプがあります。 粗い繊維状(網目状の繊維状)そして ラメラ状。これらのタイプの骨組織は構造が異なり、 物理的特性、主に細胞間物質の構造によって決まります。 骨組織には以下のものもあります 象牙質そして セメント細胞間物質の高度な石灰化(第 16 章を参照)とそれを支える機械的機能という点で骨組織に似た歯。
粗い繊維状の骨組織
粗い繊維状の骨組織 (骨網状線維筋)主に胚で発生します。 成人では、頭蓋縫合糸が伸びすぎた部位や、腱が骨に付着している部位で発生します。 ランダムに配置されたコラーゲン線維はその中で厚い束を形成しており、比較的低い顕微鏡倍率でもはっきりと見えます(図8.24)。
粗い繊維の骨組織の主物質には、細長い楕円形の形状があります 骨の空洞、または、骨細胞が存在する長い吻合細管を備えた裂孔 - 骨細胞彼らのシュートと一緒に。 骨の表面が覆われている 骨膜。
層状骨組織
層状骨組織 (textus osseus lamellaris)- 成人の体内で最も一般的な種類の骨組織。 で構成されています 骨プレート(骨層板)。後者の厚さと長さ

米。 8.24。粗繊維骨組織の構造(Yu. I. Afanasyevによる): 1 - 絡み合ったコラーゲン繊維の束。 2 - 骨細胞
それぞれ数十から数百マイクロメートルの範囲です。 それらはモノリシックではありませんが、異なる平面に配向された平行な方向を向いたコラーゲン (オセイン) フィブリルを含んでいます。 プレートの中央部分では、フィブリルは主に長手方向を持ち、周囲に沿って接線方向と横方向が追加されます。 層は剥離することがあり、1 つの層の原線維が隣接する層に続き、単一の線維性の骨基部が形成されることがあります。 さらに、骨プレートは、骨プレートに垂直に配向され、それらの間の中間層に織り込まれた個々の原線維と繊維によって貫通され、それによって層状骨組織のより大きな強度が達成されます(図8.25)。 骨格のほとんどの平らな骨や管状の骨に含まれる緻密で海綿状の物質は、この組織から作られています。
骨組織の発生(骨組織形成)
胚における骨組織の発生は 2 つの方法で行われます。1) 間葉から直接 (直接骨形成)、2) 間葉から直接発生します。 2)以前に開発された軟骨骨モデルの代わりに間葉から作成(間接骨形成)。 骨組織の胚後の発達は、生理学的および修復再生中に起こります。
骨組織には次のものが含まれます 骨芽細胞そして 破骨細胞の分化。 1 つ目 (メイン) は、幹細胞、半幹細胞 (前骨芽細胞)、骨芽細胞 (タイプ

米。 8.25。層状骨組織の構造 (Yu. I. Afanasyev による):
1 - 骨プレート。 2 - 骨細胞。 3 - 骨細胞プロセスの接触。 4 -
各骨プレート内で方向付けられたコラーゲン線維
平行
線維芽細胞)、骨細胞。 細胞の骨形成分化のプロセスは、骨形成因子(骨形成タンパク質)、組織内の酸素分圧、アルカリホスファターゼの存在などの影響を受けます。2 番目の(関連する)差異には以下が含まれます。 破骨細胞(マクロファージの一種)血液幹細胞から発生します。
幹細胞および半幹骨形成細胞は形態学的に同定されません。
骨芽細胞(ギリシャ語から。 オステオン- 骨、 爆風- rudiment) は、骨組織を形成する若い細胞です。 形成された骨では、それらは骨膜の深層と損傷後の骨組織再生の場所でのみ見られます。 それらは増殖することができ、骨の形成において、それらは発達中の骨梁の表面全体をほぼ連続した層で覆います(図8.26)。 骨芽細胞の形状は、立方体、錐体、または角張ったものなど、さまざまです。 体の大きさは約15〜20ミクロンです。 核は円形または楕円形で、多くの場合偏心して位置し、1 つまたは複数の核小体を含みます。

米。 8.26。「直接的な」骨形成:
あ- 骨形成島(図)。 b- 類骨段階(スキーム); V- 細胞間物質の骨化(スキーム); G- 平らな骨における「直接的な」骨形成(顕微鏡写真)。 1 - 間葉細胞。 2 - 毛細血管。 3 - 骨芽細胞。 4 - 類骨; 5 - 石化した細胞間物質。 6 - 骨細胞。 7 - 破骨細胞
骨芽細胞の細胞質では、顆粒小胞体、ミトコンドリア、ゴルジ複合体がよく発達しています(図8.27)。 大量の RNA とアルカリホスファターゼの高い活性が検出されます。 骨芽細胞は脂質を含む基質小胞を分泌します。

米。 8.27。骨芽細胞の構造 (Yu. I. Afanasyev による):
a - 光光学レベルで。 b- 超顕微鏡レベルで。 1 - コア。 2 - 細胞質。 3 - 顆粒小胞体。 4 - 類骨; 5 - 石灰化した骨組織
Ca 2+、アルカリホスファターゼ。有機組織マトリックスの石灰化を引き起こします。
骨細胞(ギリシャ語から。 オステオン- 骨、 サイトス- 細胞)は、分裂能力を失った骨組織の最終的な細胞の主な数です。 それらは分岐した形状、コンパクトで比較的大きな核、およびわずかに好塩基性の細胞質を持っています(図8.28)。 オルガネラは十分に発達していません。 骨細胞における中心小体の存在は確立されていません。
骨細胞は、骨細胞の輪郭に沿った骨空洞、または骨腔に存在します。 キャビティの長さは 22 ~ 55 ミクロン、幅は 6 ~ 14 ミクロンの範囲です。 骨空洞の細管は組織液で満たされており、相互および周囲と吻合しています。
骨に入る血管の眼球空間。 骨細胞と血液の間の物質の交換は、組織液を介して行われます。
破骨細胞(ギリシャ語から。 オステオン- 骨と クラストス- 断片化されています)。 これらの血行性の細胞は、石灰化した軟骨や骨を破壊する可能性があります。 それらの直径は150〜180ミクロンに達し、3〜数十個の核を含みます(図8.29)。 細胞質はわずかに好塩基性であり、場合によっては好酸性です。 破骨細胞は通常、骨梁の表面に存在します。 破骨細胞の破壊された表面に隣接する側には、微細に折り畳まれた(波形の)境界があります。 それは加水分解酵素の合成と分泌の領域です。 破骨細胞の周囲には、細胞が骨表面にしっかりと接着しているゾーンがあり、いわば酵素の作用領域を密閉します。 細胞質のこの領域は軽く、アクチンからなるマイクロフィラメントを除いて細胞小器官はほとんど含まれていません。
波形の端の上の細胞質の周縁層には、多数の小さな小胞と大きな液胞が含まれています。 破骨細胞は環境中に CO 2 を放出し、ここで見つかった酵素カルボナンヒドラーゼは酸 (H 2 CO 3 ) の生成とカルシウム化合物の溶解を促進すると考えられています。 破骨細胞にはミトコンドリアとリソソームが豊富に含まれており、その酵素(コラゲナーゼやその他のプロテアーゼ)は骨組織マトリックスのコラーゲンとプロテオグリカンを分解します。 破骨細胞が骨物質と接触する場所では、骨物質に吸収腔が形成されます。 1 つの破骨細胞は、100 個の骨芽細胞が同時に作成するのと同じ量の骨を破壊する可能性があります。 骨芽細胞の機能
sts と破骨細胞は相互に関連しており、ホルモン、プロスタグランジン、機能的負荷、ビタミンなどの関与と相関しています。
細胞間物質(substantia intercellellis)無機塩を含浸させた基本的な非晶質物質からなり、その中にコラーゲン線維が位置し、小さな束を形成しています。 それらには(最大90%)タンパク質、つまりI型コラーゲンが含まれています。 繊維は、ランダムな方向 (粗い繊維状の骨組織の場合) または厳密に配向された方向 (層状骨組織の場合) を持ちます。
骨組織の基底物質には、軟骨と比較して比較的少量のコンドロイチン硫酸が含まれていますが、カルシウムと複合体を形成して骨の有機基質に浸透するクエン酸やその他の酸が多く含まれています。 コラーゲンタンパク質に加えて、非コラーゲン性タンパク質(石灰化プロセスに関与するオステオカルシン、シアロタンパク質、オステオネクチン、オステオポンチンなど)およびグリコサミノグリカンが骨組織の主要物質に含まれています。 骨の基質には、骨の有機基質の原線維に関連して規則正しく配置されたヒドロキシアパタイトの結晶と、非晶質リン酸カルシウムが含まれています。 骨組織には 30 種類以上の微量元素 (銅、ストロンチウム、亜鉛、バリウム、マグネシウムなど) が検出され、体内の代謝プロセスで重要な役割を果たしています。 身体活動を体系的に増加させると、高度な石灰化により骨量が 10 ~ 50% 増加します。
直接的な骨組織形成。この骨形成方法は、平らな骨の形成中に粗繊維の骨組織が発達するのが特徴です。
米。 8.28。骨細胞の構造 (Yu. I. Afanasyev による):
あ- 光光学レベルで; b- 超顕微鏡レベルで。 1 - 骨細胞のプロセス。 2 - コア。 3 - 小胞体。 4 - ゴルジ複合体。 5 - ミトコンドリア。 6 - 骨細胞が位置する小腔の端に沿った類骨(石灰化されていない)骨物質

米。 8.29。破骨細胞の構造 (Yu. I. Afanasyev による):
あ- 光光学レベルで; b- 超顕微鏡レベルで。 1 - コア。 2 - 破骨細胞の波形の端。 3 - ライトゾーン。 4 - リソソーム。 5 - 細胞間物質の吸収ゾーン。 6 - 石化した物質
頭蓋骨の外皮骨などの骨。 このプロセスは主に子宮内発育の 1 か月目に観察され、一次「膜状」類骨組織の形成と、それに続く細胞間物質へのカルシウム、リン、その他の塩の含浸 (沈着) を特徴としています。 最初の段階では- 骨格形成島の形成 - 将来の骨の発達の場所では、間葉細胞の局所的な増殖と骨格形成島の血管新生が起こります。 間葉細胞は負荷ベクトルに従って配向され、骨形成前駆体である前骨芽細胞に分化します。 第二段階では島細胞は骨芽細胞に分化し、そこでコラーゲンタンパク質の生合成とその分泌が始まり、その結果、骨組織の有機マトリックスであるコラーゲン原線維を含む好酸性細胞間物質が出現します(類骨期)。 成長する繊維は細胞を押し広げますが、成長過程の助けを借りて細胞は互いに接続されたままになります。 このようにして、骨芽細胞は突起形態を獲得し、線維塊の厚さに含まれる骨細胞となり、再生能力を失う。 主要物質にはムコタンパク質 (骨ムコイド) が現れ、繊維を 1 つの強力な塊に固定します。 同時に、周囲の間葉から新しい世代の骨芽細胞が形成され、外側から骨が成長します(並置成長)。
第三段階- 細胞間物質の石灰化(塩の含浸)。 この場合、骨芽細胞はアルカリホスファターゼという酵素を分泌し、末梢血に含まれるグリセロリン酸を炭水化物化合物(糖)とリン酸に分解します。 後者は、主物質および繊維に沈着するカルシウム塩と反応し、最初は非晶質を形成するカルシウム化合物の形で反応します。
[Ca 3 (PO 4) 2] が堆積し、続いてヒドロキシアパタイト [Ca 10 (PO 4) 6 (OH) 2] の結晶がそこから形成されます。
石灰化のメディエーターの 1 つは、 オステオネクチン- カルシウム塩とリン塩をコラーゲンに選択的に結合する糖タンパク質。 石灰化の結果、骨の横棒、つまり梁が形成されます。 次に、これらの横木から枝分かれし、相互接続して広いネットワークを形成します。 クロスバー間の空間は、血管が通過する緩い線維性結合組織によって占められていることが判明した。
組織形成が完了するまでに、骨胚の周囲に沿った胎児結合組織に多数の線維と骨形成細胞が現れます。 骨梁に直接隣接するこの線維性結合組織の一部は、 骨膜、栄養と骨の再生をもたらします。 このような骨は、胚発生の段階で出現し、粗い繊維状の骨組織の支柱からなり、一次海綿骨と呼ばれます。 発達の後期段階では、成人の二次海綿骨に置き換わりますが、層状骨組織から構築されている点で最初の海綿骨とは異なります (骨形成の第 4 段階)。 層状骨組織の発達は、骨の個々の部分の破壊および粗繊維骨の厚さへの血管の内方成長のプロセスと密接に関連しています。 破骨細胞は、胎児の骨形成中と出生後の両方でこのプロセスに参加します(図8.29を参照)。 骨芽細胞の層が血管の周囲に形成され、その後、新しい同心の骨板が現れます。 各層のコラーゲン線維は平行ですが、前の層の繊維に対してある角度をなして配向されています。 したがって、骨の円柱が血管の周囲に形成され、一方が他方に挿入されます(一次骨)。 骨が出現した瞬間から、粗い線維性骨組織は発達を停止し、層状骨組織に置き換えられます。 骨膜の側からは、外側を取り囲む(一般、一般)プレートが形成され、骨全体を外側から覆います。 これが平らな骨が発達する方法です。 その後、胎児期に形成された骨は再構築を受けます。一次骨骨が破壊され、新しい世代の骨骨が発達します。 この骨の再構築は事実上生涯を通じて続きます。
軟骨組織とは異なり、骨は常に既存の組織に新しい組織を適用することによって成長します。 並置、そして、骨格形成膵島細胞の分化には最適な血液供給が必要です。
間接的な骨組織形成。胚発生の 2 か月目に、将来の管状骨の場所で間葉から軟骨の基礎が形成され、これは非常に急速に将来の骨 (軟骨モデル) の形をとります。 原始体は、軟骨膜で覆われた胎児硝子軟骨で構成されています(図8.30)。 しばらくの間、軟骨膜の側面に形成された細胞と内部領域の細胞の増殖の両方により成長します。

米。 8.30。間接的(軟骨性)骨形成。 骨および軟骨周囲骨カフの軟骨モデルの形成 (Yu. I. Afanasyev による):
a-d - 骨形成の段階。 1 - 管状骨の一次軟骨モデル。 2 - 軟骨膜; 3 - 軟骨組織。 4 - 軟骨周囲骨カフ。 5 - 骨膜。 6 - 軟骨細胞の列。 7 - 小胞細胞のゾーン。 8 - 間葉は破骨細胞 (9) と毛細血管 (10) の分化とともに軟骨に成長します。 11 - 骨芽細胞。 12 - 軟骨内に形成された骨組織。 13 - 骨端の骨化点
軟骨の代わりの骨の発達、つまり間接的な骨形成は、骨幹の領域で始まります(軟骨膜骨化)。 教育 軟骨周囲骨リング(カフ)骨幹の中央部分に隣接する軟骨膜での分化を伴う血管の増殖が先行して、骨芽細胞が最初にカフを形成します。 粗い繊維状の骨組織(一次骨化中心)、その後交換 ラメラ状。
骨のカフが形成されると、軟骨の栄養が破壊されます。 その結果、軟骨原基の骨幹部分の中心に異栄養性変化が発生します。 軟骨細胞は空胞化し、その核が濃縮され、いわゆる小胞性軟骨細胞が形成されます。 この場所での軟骨の成長は止まります。 軟骨周囲骨輪の拡大には、軟骨破壊領域の増加と破骨細胞の出現が伴い、管状骨モデル内に成長する血管と骨芽細胞の通路が確保されます (図 8.30 を参照)。 これにより、軟骨内骨化の病巣(二次骨化中心)が出現します。 骨幹の隣接する未変化の遠位部分の継続的な成長により、骨端と骨幹の境界にある軟骨細胞が柱状に収集され、その方向は将来の骨の長軸と一致します。 したがって、軟骨細胞の柱には、2つの反対方向のプロセス、つまり骨幹の遠位部分での再生と成長、およびその近位部分でのジストロフィープロセスが存在します。 同時に、腫れた細胞の間にミネラル塩が沈着し、重度の好塩基球増加症の出現と軟骨の脆弱化を引き起こします。
血管網が成長して骨芽細胞が出現した瞬間から、軟骨膜が再構築され、 骨膜。その後、周囲の間葉、骨形成細胞、破骨細胞を含む血管が骨カフの開口部を通って成長し、石灰化軟骨と接触します(図8.30を参照)。 破骨細胞によって分泌される酵素の影響下で、石灰化した細胞間物質の溶解(軟骨溶解)が起こります。 骨幹軟骨が破壊され、その中に細長い空間が現れ、そこに骨芽細胞が「定着」し、軟骨の石灰化細胞間物質の残りの領域の表面に骨組織を形成します。
一次、または骨幹の骨化中心。軟骨の原始内部での骨形成のプロセスは、軟骨内骨化と呼ばれます(ギリシャ語。 エンドン- 内部)。
軟骨内骨の発達と同時に、破骨細胞による破壊の兆候が現れます。 軟骨内骨組織の破壊により、さらに大きな空洞と空間(吸収空洞)が形成され、最終的には 骨髄腔。ここに浸透する間葉から骨髄間質が形成され、そこに血液および結合組織の幹細胞が定着します。 同時に、骨膜の側面から骨幹の周囲に沿って、骨組織の新しい横棒がますます出現します。 骨組織は、骨端に向かって長さが増加し、厚さが増加して、緻密な骨の層を形成します。
骨膜骨の組織化は、軟骨内骨組織の組織化とは異なる方法で進行します。 崩壊した粗繊維骨の代わりに、骨原始の長軸に沿ってここに侵入した血管の周囲に、平行に配向した薄いコラーゲン繊維と接着細胞間物質からなる同心円状のプレートが形成され始めます。 こうして彼らは生まれるのです 一次骨。それらの内腔は広く、プレートの境界は輪郭がはっきりしていません。 第一世代のオステオンの出現に続いて、 外部包囲(一般)プレート、骨幹領域の骨の周囲。 骨幹に続いて、骨端に骨化中心が現れます。 これには、まず軟骨細胞の分化、肥大が起こり、続いて栄養の低下、細胞間物質の変性、石灰化が起こります。 その後、上記と同様の骨化のプロセスが観察されます。 骨化は、血管の骨端への内方成長を伴います。
骨幹と骨端の間の中間領域では、軟骨組織が保存されています。 骨幹端軟骨、骨の長さが成長するゾーンです。
臓器としての管状骨の組織構造
器官としての管状骨は、結節を除いて主に層状骨組織から構成されています。 骨の外側は、一種の硝子軟骨で覆われている骨端の関節面を除いて、骨膜で覆われています。

米。 8.31。管状骨の構造 (V. G. Eliseev、Yu. I. Afanasyev、E. F.kotovsky による):
あ- 骨膜; b- 緻密な骨物質。 V- 内骨; G- 骨髄腔。 1 - 外側の共通プレートの層。 2 - オステオン; 3 - オステオンチャネル。 4 - インサートプレート。 5 - 内部共通プレートの層。 6 - 海綿状物質の骨梁。 7 - 骨膜の繊維層。 8 - 骨膜の血管。 9 - 穿孔チャネル。 10 - 骨細胞
骨膜、または 骨膜。骨膜には、外側 (繊維) と内側 (細胞) の 2 つの層があります。 外層は主に線維性結合組織で形成されています(図8.31、8.32)。 内層には、形成層細胞、前骨芽細胞、さまざまな分化度の骨芽細胞など、多数の細胞が含まれています。 紡錘形の形成層細胞は、少量の細胞質と適度に発達した合成装置を持っています。 前骨芽細胞は、グリコサミノグリカンを合成できる、活発に増殖する楕円形の細胞です。 骨芽細胞は、よく発達したタンパク質合成 (コラーゲン) 装置を特徴としています。 骨に栄養を供給する血管と神経は骨膜を通過します。
骨膜は骨を周囲の組織と接続し、その栄養、発達、成長、再生に関与します。
骨幹の構造。骨幹を形成する緻密な物質は骨プレートで構成され、その厚さは 4 ~ 12 ~ 15 ミクロンの範囲です。 骨プレートは一定の順序で配置されており、

米。 8.32。骨膜 (Yu. I. Afanasyev による):
1 - 外側(繊維)層。 2 - 内(細胞)層。 3 - 骨形成細胞。 4 - 骨組織
複雑なシステムを形成しています。 骨幹には 3 つの層があります。 外側周囲を囲む(一般、一般)プレートの層、 平均、血管の周りに同心円状に層状に重なった骨プレートによって形成されます - オステオンであり、オステオン層と呼ばれます (図8.33)。 インテリア周囲を取り囲む(共通の)プレートの層。
外側のプレートは骨幹の周りに完全なリングを形成せず、後続のプレートの層によって表面に重ねられます。 内部プレートは、緻密な骨物質が髄腔に直接接する場所でのみよく発達します。 緻密物質が海綿状物質に移行するのと同じ場所で、その内部の共通プレートは海綿状物質の小柱のプレートに続いています。
外板に穴が開いています 穿孔(フォルクマン)チャネル、そこを通って血管が骨膜から骨に進入します。 骨膜から、コラーゲン線維がさまざまな角度で骨に浸透します。 これらの繊維はと呼ばれます 穿孔(シャーピー)繊維。ほとんどの場合、それらは共通層の外層でのみ分岐しますが、中間の骨層に浸透することもありますが、骨層には決して入りません。
中層には骨があります 同心(骨)プレートオステオンを形成します。 オステオンの間に位置する 中間(インサート)プレート。骨プレートの厚さと長さは数十から数百マイクロメートルの範囲にあります。 オステオン(Haversian システム) は、管状骨の緻密な物質の構造単位です (図 8.31、図 8.33 を参照)。 それらは、あたかも互いに挿入されているかのように、同心の骨プレートからなる円筒形の形成物です。 骨プレートと骨プレートの間には、骨細胞の本体とその突起があり、骨の細胞間物質に浸されています。 各オステオンは、基質によって形成されるセメント(融合)線によって隣接するオステオンから区切られています。 中央にある

米。 8.33。オステオン:
あ- 顕微鏡写真(シュモール法を使用した染色)。 b- 骨片の走査型電子顕微鏡検査(O. V. Slesarev による作成)。 1 - オステオンチャネル; 2 - 骨細胞(空洞 - b); 3 - 骨プレート
オステオンチャネルには、結合組織および骨形成細胞を伴う血管が含まれています。
長骨の骨幹では、骨は主に長軸と平行に位置しています。 骨管は互いに吻合し、吻合部位では隣接するプレートの方向が変わります (図 8.31 を参照)。 このようなチャネルは次のように呼ばれます。 穿孔、または 栄養補給。骨管に位置する血管は、相互に連絡し、また骨髄および骨膜の血管とも連絡します。 骨幹の大部分は、管状骨の緻密な物質で構成されています。 直径の内面に
髄腔に隣接する骨層、層状骨組織は海綿骨の骨横棒を形成します。 管状骨の骨幹の空洞は骨髄で満たされています。
骨内膜 -骨を髄腔側から覆う膜。 形成された骨表面の骨内膜では、石灰化した骨物質の外縁にオスミウム親和性の線が認められます。 類骨層は、非晶質物質、コラーゲン原線維と骨芽細胞、毛細血管と神経終末からなり、骨内膜を骨髄の要素から漠然と分離する細胞の層です。 骨内膜の厚さは 1 ~ 2 ミクロンを超えますが、骨膜の厚さよりは薄いです。
骨形成が活発な領域では、骨芽細胞とその前駆体の合成活性の増加による類骨層により、骨内膜の厚さが 10 ~ 20 倍に増加します。 骨の再構築中に、破骨細胞が骨内膜で見つかります。 老化した骨の骨内膜では、骨芽細胞と前駆細胞の数が減少しますが、破骨細胞の活動が増加し、緻密層が薄くなり、海綿骨が再構築されます。
骨内膜と骨膜の間には、骨組織のラクナ管状系により、体液とミネラルの特定の微小循環が存在します。
骨組織の血管新生。血管は骨膜の内層に密なネットワークを形成します。 ここから細い動脈の枝が発生し、栄養開口部を通って骨に血液を供給し、骨髄を貫通して骨髄に栄養を供給する毛細血管網の形成に関与します。 リンパ管は主に骨膜の外層に位置しています。
骨組織の神経支配。骨膜では、有髄神経線維と無髄神経線維が神経叢を形成します。 繊維の一部は血管に付随し、血管とともに栄養開口部を通って同名の管に侵入し、次に骨管に入り、骨髄に到達します。 線維の別の部分は自由神経枝を備えた骨膜で終わり、またカプセル化された体の形成に関与します。
管状骨の成長。骨の成長は非常に長いプロセスです。 ヒトでは胎児期の初期から発症し、平均して20歳までに終了します。 成長期全体を通じて、骨は長さと幅の両方が増加します。 管状骨の長さの成長は、2つの相反する組織形成過程が現れる中骨端軟骨成長板の存在によって確実に行われます。
1 つは骨端板の破壊で、もう 1 つはその逆で、新しい細胞の形成による軟骨組織の継続的な補充です。 しかし、時間の経過とともに、細胞破壊のプロセスが新生物のプロセスよりも優先され始め、その結果、軟骨板が薄くなり、消失します。 骨の長さの成長が止まります。
中骨端軟骨には、 国境ゾーン、ゾーン 円柱状細胞そしてゾーン 小胞細胞。近くにある国境地帯
骨端は、円形および楕円形の細胞と、軟骨板と骨端の骨の間の接続を提供する単一の同質遺伝子グループから構成されます。 骨と軟骨の間の空洞には、軟骨板のより深い領域の細胞に栄養を与える毛細血管があります。 円柱状細胞ゾーンには、骨の長手方向軸に沿って柱を形成し、その成長と長さをもたらす活発に増殖する細胞が含まれています。 カラムの近位端は、成熟し分化している軟骨細胞で構成されています。 グリコーゲンとアルカリホスファターゼが豊富に含まれています。 これらのゾーンはどちらも、骨化と骨の成長のプロセスに影響を与えるホルモンやその他の要因の作用に対して最も反応性が高くなります。 小胞細胞ゾーンは、軟骨細胞の水和と破壊、その後の軟骨内骨化によって特徴付けられます。 このゾーンの遠位部分は骨幹に隣接しており、そこから毛細血管と骨形成細胞が骨幹に侵入します。 軟骨内骨の長手方向の柱は、本質的に骨が形成される骨管です。
その後、骨幹と骨端の骨化中心が結合し、骨の長さの成長が終了します。
管状骨の幅の増加は骨膜によるものです。 骨膜の側では、層状骨組織の同心円層が非常に早期に形成され始めます。 これ 同位的な成長は骨の形成が完了するまで続きます。 出生直後の骨の数は少ないですが、25歳までに手足の長骨の骨の数が大幅に増加します。
再生。骨組織の生理学的再生は、骨膜 - 骨内膜の骨形成細胞および骨管内の骨形成細胞によってゆっくりと起こります。 外傷後の骨組織の再生は、骨折した骨の端が相互にずれていない場合に、より良好に進行します。 骨形成のプロセスには結合組織カルスの形成が先行し、その厚さに軟骨島が形成される可能性があります(図8.34)。 この場合の骨化は、二次(間接的)骨形成の種類に応じて発生します。 最適な組織酸素化、骨折端の適切な再配置および固定の条件下では、カルスを形成することなく再生が起こります。 しかし、骨芽細胞が骨を作り始める前に、破骨細胞は骨の接合端の間に小さな隙間を形成します。 この生物学的パターンは、外傷専門医が再生の全期間を通じて癒合した骨を徐々に伸ばすための装置を使用するための基礎となっています。
骨のリモデリングとその構造に影響を与える要因
骨組織では、機能的負荷や外部および内部環境のその他の要因によって、生涯を通じて相互に関連した破壊と創造のプロセスが発生します。 骨の再構築は常に、一次骨の破壊と、破壊部位と骨膜の側面の両方での新しい骨の同時形成と関連しています。 破骨細胞の影響で活性化

米。 8.34。外傷後の管状骨の再生: あ- 損傷の局在化。 神- 縮小した骨をしっかりと固定しない、連続的な再生段階 (b 1、 1で- 断片); d- 断片の固定後の再生。 1 - 骨膜。 2 - 粗い繊維の骨組織で作られたクロスバー。 3 - 結合組織は軟骨組織の島とともに再生します。 4 - 骨は粗い線維性骨組織から再生します。 5 - 融合ライン (R.V. Krstic による、修正あり)
さまざまな要因により、骨の骨板が破壊され、その場所に空洞が形成されます。 このプロセスはと呼ばれます 吸収(緯度から。 吸収不全- 骨組織の吸収)。 骨芽細胞は、残りの血管の周りの結果として生じる空洞に現れ、互いに同心円状に積層された新しいプレートの構築が始まります。 これがオステオンの第二次世代の発生方法です。 オステオンの間には、前世代の破壊されたオステオンの残骸があります。 骨の再構築のプロセスは、骨の成長が終了した後でも止まりません。
骨組織の再構築に影響を与える要因の中で、いわゆる圧電効果が重要な役割を果たします。 骨プレートを曲げると、凹面と凸面の間に一定の電位差が現れることが分かりました。 最初のものはマイナスに帯電しており、
そして2番目はポジティブです。 負に帯電した表面では、骨芽細胞の活性化と骨組織の付加的な新たな形成のプロセスが常に観察されますが、正に帯電した表面では、逆に、破骨細胞の助けによるその吸収が観察されます。 人為的に電位差を作成しても同じ結果が得られます (図 8.35)。 ゼロ電位、骨組織への物理的ストレスの欠如(長時間の固定、無重力状態など)は、破骨細胞の機能の増加と塩の排出を引き起こします。
骨組織と骨の構造は、ビタミン(C、A、D)、甲状腺、副甲状腺、その他の内分泌腺のホルモンの影響を受けます。
特に、体内のビタミンCの量が不十分な場合(壊血病など)、コラーゲン線維の形成が抑制され、骨芽細胞の活性が弱まり、ホスファターゼ活性が低下するため、事実上、ビタミンCの摂取が停止します。骨組織の有機的基盤の形成の阻害による骨の成長。 ビタミン D 欠乏症(くる病)では、有機骨基質の完全な石灰化が起こらず、骨が軟化します(骨軟化症)。 ビタミン A は骨の成長をサポートしますが、このビタミンが過剰になると、骨の成長領域である破骨細胞による中骨端軟骨の破壊が増加し、骨の伸長が遅くなります。
副甲状腺ホルモンであるパラチリンが過剰になると、破骨細胞の活動の増加と骨吸収が観察されます。 甲状腺のC細胞によって産生されるチロカルシトニンは、正反対の働きをし、このホルモンの受容体を持つ破骨細胞の機能を低下させます。 甲状腺の機能が低下すると、骨芽細胞の活動が抑制され、骨化プロセスが阻害されるため、長い管状骨の成長が遅くなります。 この場合の骨の再生は弱く不完全に起こります。 精巣の発育不全または思春期前の去勢の場合、中骨端板の骨化が遅れ、その結果、そのような人の腕と脚が不釣り合いに長くなります。 閉経後にエストロゲンが不足すると、女性は骨粗鬆症を発症することがあります。 思春期早期になると、骨幹と骨端の骨の早期癒合が原因で成長が停止すると予想されます。 骨の成長における一定の積極的な役割は、下垂体腺の成長ホルモンによって演じられ、若い(青年期)年齢では比例した骨格の発達を刺激し、成人では不均衡(先端巨大症)の骨格の発達を刺激します。
年齢は変わります。結合組織は、年齢とともに構造、量、化学組成が変化します。 年齢とともに、結合組織形成の総質量と骨骨格の成長が増加します。 多くの種類の結合組織構造では、コラーゲンとグリコサミノグリカンの種類の比率が変化します。 特に、硫酸化化合物がより多く含まれています。
骨の接続
2 つのボーンを接続できる 継続的な(癒合症、癒合症、癒合症)および 間欠(関節)。
継続的な接続- 高密度の線維性結合組織の助けを借りた接続。その束が穿孔線維の形で骨に導入されます。

米。 8.35。圧電効果 (本文中の説明): I - 骨梁の構造組織の図。 II - 骨梁の形状が変化するときの破骨細胞と骨芽細胞の活性化。 III - 電位差の人為的生成 (Yu. I. Afanasyev による)
新しい生地。 そのような接続の例は、頭蓋骨の頭頂骨の縫合、橈骨と尺骨の間の結合組織膜です。
シンコンドローゼ (結合)- 椎間板などの軟骨によって形成される接続。 それらは外側の線維輪と髄核と呼ばれる内側の部分で構成されています。 これらの部分は両方とも漠然と分離されており、気づかれないうちに相互に変化します。 髄核は椎間板の内側領域に位置しています。 さまざまな年齢層で、それは 異なる構造。 2歳までの年齢では、個々の細胞のみを含む均質な内容物を含む空洞になります。 その後の人生では、この空洞は別々の部屋に分割されます。 6~8歳になると、髄核にコラーゲン線維や軟骨細胞の出現と数の増加が認められます。 15 歳から、線維と軟骨細胞の成長はさらに激化し、20 ~ 23 歳になると、髄核は線維軟骨の特徴的な外観になります。 別のより緊密な接続の例としては、恥骨結合が挙げられます。 結合軟骨には、中骨端軟骨の助けを借りた骨端と骨幹の接続も含まれます。
癒合症- 骨盤骨など、線維性結合組織のない骨の堅い関節。
断続的な関節、または関節 (関節症) は、軟骨で覆われた関節面と、場合によっては軟骨の中間半月板と関節包で覆われた関節面で構成されています。 関節包は外側の繊維層と内側の滑膜層で構成されています。 後者は、血管、リンパ管、神経線維、終末を含む、特に分化した結合組織の層として理解されています。 他の間葉系誘導体には珍しく、この結合組織の境界線の位置、関節の運動機能の関与による一定の伸張、変位、圧力が成長とその構造的特徴を決定します。
哺乳動物とヒトの滑膜では、2 つの線維性コラーゲン弾性層 (表層と深層) と腔を裏打ちする被覆層が区別されます (図 8.20 を参照)。 層間に明確な境界はありません。 大きな関節では、線維性被膜に隣接する脂肪組織が豊富な滑膜下層があります。 表層のコラーゲンと弾性繊維は関節の長軸方向に配向しています。 深層では、それらは表層の繊維に対してある角度で位置しています。
滑膜の被覆層は細胞、つまり滑膜細胞で構成されています。 マクロファージ滑膜細胞と滑膜線維芽細胞があり、これらは滑液の特定の成分であるヒアルロン酸を生成および分泌する能力を持っています。
血管は下層の組織から滑膜を貫通し、滑膜細胞の直下に位置する外皮層を含む滑膜の厚さ全体に分布しています。 したがって、滑膜腔は、結合組織の主要物質である細胞と毛細血管自体の内皮によってのみ血流から分離されています。 滑膜の毛細血管の内皮は、窓窓と貪食能力によって特徴付けられます。 毛細リンパ管は常に表層線維層内の毛細血管よりも深いところに位置しています。
滑膜は、求心性および遠心性(交感神経)性質の線維によって豊富に神経支配されています。
コントロールの質問
1. 結合組織の発達の源としての間葉: 間葉の分岐分化の概念、間葉の形態学的特徴。
2.特別な特性を持つ結合組織:分類、体内の地形、構造、機能。
3. 疎性結合組織の細胞の違い: 発生の源、構造、機能、生理学的および修復再生への関与。
4. 軟骨組織:分類、地形、構造、機能、再生。
5. 骨組織:直接的および間接的な骨形成、構造、細胞分化組成、再生。
組織学、発生学、細胞学:教科書 / Yu. I. Afanasyev、N. A. ユリナ、E. F. コトフスキー、他 - 第 6 版、改訂。 そして追加の - 2012年。 - 800ページ。 : 病気。