成熟(RNAプロセシング)。 加工、つなぎ合わせ。 遺伝情報を実現する過程における RNA の役割 真核生物における一次転写物の処理
原核生物のタンパク質をコードする遺伝子の転写によって生成される一次転写物は、その後の修飾やプロセシングを行わずに mRNA として機能します。 実際、mRNA の翻訳は、転写産物の 3 インチ末端の合成が完了する前に始まることがよくあります。rRNA 分子と tRNA 分子では、まったく異なる状況が観察されます。この場合、rRNA または tRNA 遺伝子のクラスター、またはこれらの断片が散在している場合もあります。遺伝子は転写されて単一の RNA 鎖を形成することがよくあります。これらの遺伝子の転写は常に特定のプロモーターで始まり、特定のターミネーターで終了しますが、成熟した機能的形態を生成するには、一次 RNA 転写物の特定の切断と修飾が発生する必要があります。このような分子イベントは集合的に呼ばれます。 rRNA と tRNA のプロセシングのメカニズムと、それを実行する酵素については、最も詳しく研究されています。 大腸菌、転写後の RNA プロセシングの特徴を説明するために、このシステムを使用します。 真核生物の RNA に対する同様の修飾。 この場合、rRNA と tRNA のプロセシングに加えて、mRNA の形成を伴う転写物の成熟のためのより複雑なシステムが使用されます。
A. rRNAおよびtRNAをコードする遺伝子群
ゲノム内 大腸菌 rRNAをコードする7つの個別の転写単位が同定され、マッピングされています。 各転写単位は、約 5000 ヌクレオチドで構成され、5S、16S、および 23S rRNA のコード配列を 1 コピーずつ含む RNA 分子です。 この領域での転写は、16S -> 23S -> 5S の方向に発生します。 これら 3 つの rRNA コード配列に加えて、転写物にはさまざまな長さのインサートと 1 つ以上の tRNA 遺伝子のコピーが含まれています。 スペーサーは rRNA の配列の前、間、または後に配置することができ、tRNA 遺伝子は通常、点在するスペーサー セグメントまたは 3 インチ末端のスペーサー セグメント内にあります。機能的に成熟した RNA 分子を形成するには、そのような転写物のプロセシングが発生する必要があります。プロセシングの前または途中で、スペーサー、rRNA、tRNA 遺伝子の特定の塩基。
b. rRNA-tRNA共転写物の切断
tRNA または 16S-、23S-、または 58-rRNA 配列を含むフラグメントへの一次転写物の最初の切断は、エンドヌクレアーゼ RNase III によって実行されます。 その標的は、各rRNAセグメントに隣接する配列内の分子内塩基対形成によって形成される短いRNA二重鎖です。 例えば、16S-pRNA配列に隣接するスペーサー領域内の相補的領域はヘアピンのステムを形成し、そのループには16S-pRNA配列が含まれる。 23S-rRNA 配列と 5S-rRNA 配列も同様のヘアピンを形成します。 RNase III は二本鎖ステムに切断を導入し、5"-リン酸末端と 3"-ヒドロキシル末端を持つ短いスペーサー領域に隣接した、1 つまたは別の rRNA の配列を含む RNA 鎖を形成します。 次に、おそらく tRNA プロセシングの最終段階を触媒する同じ RNA エキソヌクレアーゼによって、スペーサー配列の余分なヌクレオチドが除去されます。 原則として、酵素による切断が起こるには、ヘアピンを形成するヌクレオチド配列のみが転写されなければなりません。 ただし、エンドヌクレアーゼ III によって認識される RNA 転写物全体の正しいフォールディングには明らかに、リボソームまたはその他のタンパク質が必要であるため、プロセシングは一次転写物全体の合成の完了後にのみ発生します。 複数遺伝子の転写物から放出された tRNA セグメントのプロセシングは、単一遺伝子の転写単位からの tRNA のプロセシングと同じ方法で実行されます。
V. より大きな転写物からの成熟tRNAの形成
一部の tRNA コード遺伝子は rRNA 転写単位内に位置し、rRNA 遺伝子と一緒に発現しますが、大部分の tRNA 遺伝子は単一遺伝子またはクラスターで表されます。 同じ遺伝子の複数の繰り返しを含むクラスターもあれば、異なる無関係な tRNA 遺伝子を含むクラスターもあります。 場合によっては、各クラスターは 1 つの大きな RNA 分子として転写され、これが処理されて成熟 tRNA 断片が順次放出されます。 成熟した機能的 tRNA を形成するには、塩基の特異的な修飾と 3'-CCA 末端の 1、2、または 3 つすべてのヌクレオチドの付加が明らかに起こらなければなりません。
一次転写産物に 1 つ以上の tRNA 配列が含まれているかどうか、またはこれらの配列が rRNA スペーサー領域に埋め込まれているかどうかに関係なく、すべての tRNA の 5 インチ末端は RNase P と呼ばれる単一のエンドヌクレアーゼによって形成されます。RNase P は特徴的な折り畳み構造を認識すると思われます。前駆体ポリヌクレオチド内のtRNAの末端を切断し、成熟tRNA配列の5”末端の前に位置するリーダーまたはスペーサー配列を切断する。 tRNA の 3 インチ末端は、いくつかの活性によって形成されます。まだ未確認のエンドヌクレアーゼが、成熟 tRNA の 3 インチ末端が位置するヘアピン部位で前駆体を切断し、その後、別のエンドヌクレアーゼ RNase D が正しい 3 インチ末端の形成を完了します。場合によっては、エキソヌクレアーゼによる切断は成熟 tRNA の 3'-CCA 末端で正確に停止しますが、他の場合には、エキソヌクレアーゼの作用下で、プライマーとして機能する末端が形成され、tRNA のヌクレオチジルが形成されます。トランスフェラーゼは、1 つまたは複数の不変末端ヌクレオチドを追加します。
RNase P の際立った特徴は、その切断部位が tRNA 分子の正しいフォールディングの結果として形成されることです。 このフォールディングの破壊をもたらさないヌクレオチド配列の変化は、5 末端のプロセシングには影響しません。RNase P のもう 1 つの珍しい特性は、タンパク質と RNA で構成されていることです。この RNA は、377 ヌクレオチドと 377 ヌクレオチドの特異的な配列を持っています。 RNA はそれ自体、わずかに大きな遺伝子から RNA ポリメラーゼによって転写され、その後、成熟分子のサイズにプロセシングを受けます。この RNA の驚くべき特徴は、単独でリボ核タンパク質全体と同じエンドヌクレアーゼ反応を触媒できることであり、このタンパク質には存在しません。したがって、エンドヌクレアーゼ活性は RNA 自体に固有である可能性があり、タンパク質は RNA 構造を最大限に活性な構成に維持する必要があると考えられます。
成熟した tRNA は特徴的な立体構造を持っているだけでなく、修飾されたヌクレオチドも含んでいます。 これらの修飾の多くは、tRNA の生理学的機能の一部に不可欠であると考えられます。 現在、膨大な数の修飾反応を触媒する酵素群のうち、特徴づけられているのはほんのわずかです。 ただし、修飾は主に前駆体 RNA の段階と完全に処理された tRNA で発生することは明らかです。 このような修飾酵素は、その異常な配列特異性により特に興味深いものとなっています。たとえば、個々のウラシル残基のみがチオウラシルに変換され、チミンにメチル化され、またはジヒドロウラシルに還元されます。 さらに不思議なのは、ウラシルとリボースの通常の結合が修飾されるとプソイドウリジル酸が形成されることです。
mRNA のキャッピングとポリアデニル化は、 処理 (転写後修飾)。
キャッピング:
すべての真核生物の mRNA は、プロセシング中に 5 インチ末端に残基が追加されます。 7-メチルグアノシン教育を受けて ユニークな5インチ×5インチのホスホジエステル結合。 この追加のヌクレオチドは次のように呼ばれます。 キャップまたは キャップ。
キャップ機能 :
1. RNAをエキソヌクレアーゼから保護します
2. mRNA 分子のリボソームへの結合を助けます。
ポリアデニル化:
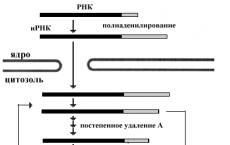
3"末端も転写完了直後に修飾されます。特殊な酵素が ポリアデニレートポリメラーゼ 20 ~ 250 個のアデニル酸残基 (ポリ(A)) を各 RNA 転写物の 3 インチ末端に結合します。ポリアデニレート ポリメラーゼは特定の配列を認識します。 あああああ、一次転写産物から 11 ~ 30 ヌクレオチドの小さなフラグメントを切断し、ポリ (A) 配列を結合します。 このような「尾部」がその後の RNA のプロセシングと核からの成熟 mRNA 分子の輸送に寄与していることは一般に受け入れられています。

mRNA が翻訳プロセスに関与するにつれて、ポリ A フラグメントの長さは減少します。 30 個のアデニル ヌクレオチドが安定性にとって重要であると考えられています。
RNA ポリメラーゼ II の核転写産物のセット全体は、次のように知られています。 不均一な核RNA(hnRNA)。
3 つのクラスの RNA はすべて、以下を含む遺伝子から転写されます。 イントロン(情報のない領域) とエクソン(情報を伝える DNA の部分)。 DNA イントロンによってコードされる配列は、RNA が生物学的に活性になる前に、一次転写物から除去する必要があります。 イントロン配列のコピーを削除するプロセスは、 RNAスプライシング.

RNAスプライシングは触媒される タンパク質とRNAの複合体、 として知られている 「小さな核リボ核タンパク質粒子」(snRNP、英語の小さな核リボ核粒子、 snRNPこのような触媒性 RNA は次のように呼ばれます。 リボザイム。
イントロンの機能:
細胞ゲノムの機能的に活性な部分を化学的または物理的(放射線)因子の有害な影響から保護する
· いわゆる 代替スプライシング遺伝子の数を増やさずにゲノムの遺伝的多様性を増加させます。

代替スプライシング:
スプライシング中に 1 つの転写産物のエクソンの分布が変化した結果、異なる RNA が生成され、その結果、異なるタンパク質が生成されます。
40 を超える遺伝子がすでに知られており、その転写物は選択的スプライシングの対象となります。 たとえば、カルシトニン遺伝子の転写物は、選択的スプライシングの結果として RNA を生成します。RNA は、カルシトニン (甲状腺内) または味覚を司る特定のタンパク質 (脳内) の合成の鋳型として機能します。 α-トロポミオシン遺伝子転写物はさらに複雑な選択的スプライシングを受けます。 単一の転写産物に由来する少なくとも 8 つの異なるトロポミオシン mRNA が同定されています (図を参照)

33. タンパク質生合成の一般的なスキーム - 必要な前提条件:
情報の流れは、情報を伝達するための仕組みです(分子生物学のセントラルドグマ)。 DNA の複製と転写 - 酵素、メカニズム。 逆転写、リバーターゼの役割。 mRNAのプロセシングとスプライシング。 遺伝暗号、コドン、アンチコドンの特徴。
タンパク質生合成と他の分子の生合成の違い:
・マトリックス中のモノマー数と反応生成物中のモノマー数(4ヌクレオチド - 20アミノ酸)の間には対応関係がありません。
・mRNA(鋳型)とタンパク質(生成物)のペプチド鎖の間には相補性がありません。
タンパク質生合成の一般的なスキーム - 必要な前提条件:
· 情報の流れ(DNAからRNA、そしてタンパク質への情報の伝達)
· プラスチックフロー(アミノ酸、mRNA、tRNA、酵素)
· エネルギーの流れ(マクロエネルギーATP、GTP、UTP、CTP)
合成直後、一次 RNA 転写物はさまざまな理由からまだ活性を持たず、「未成熟」であり、その後プロセシングと呼ばれる多くの変化を受けます。 真核生物では、すべての種類のプレ RNA がプロセシングされますが、原核生物では、rRNA と tRNA 前駆体のみがプロセシングされます。
メッセンジャーRNA前駆体のプロセシング
タンパク質に関する情報を運ぶ DNA セクションを転写すると、mRNA よりもサイズがはるかに大きい、不均一な核 RNA が形成されます。 実際のところ、遺伝子のモザイク構造により、これらの異種 RNA には情報 (エクソン) 領域と非情報 (イントロン) 領域が含まれています。
1. スプライシング スプライス- 突合せ接着)は、参加者が参加する特別なプロセスです。 核内低分子RNAイントロンは除去され、エクソンは保持されます。
スプライシング イベントのシーケンス
2. キャッピング キャップ– ヘッダー) – 転写中に発生します。 このプロセスは、5 インチ炭素 N 7 -メチルグアノシンをプレ mRNA の末端ヌクレオチドの 5 インチ三リン酸に付加することから構成されます。
「キャップ」は、mRNA のリボソームへの結合や翻訳の開始だけでなく、5 末端から作用するエキソヌクレアーゼから RNA 分子を保護するためにも必要です。
3. ポリアデニル化– ATP 分子を使用するポリアデニル酸ポリメラーゼの助けにより、100 ~ 200 個のアデニル ヌクレオチドが RNA の 3 インチ末端に結合し、ポリアデニル フラグメントであるポリ (A) テールを形成します。ポリ (A) テールは保護するために必要です。エキソヌクレアーゼからの RNA 分子は 3 インチ末端から作用します。

処理後のメッセンジャー RNA の模式図
リボソームRNA前駆体のプロセシング
rRNA 前駆体は、成熟 rRNA と比較して大きな分子です。 それらの成熟は、プレリボソーム RNA をより小さな形態に切断することにつながり、これはリボソームの形成に直接関与します。 真核生物には4種類のrRNAがあります- 5S-、5.8S-、18S-、および 28S-rRNA。 この場合、5S rRNA は個別に合成され、大きなプレリボソーム 45S RNA は特異的な酵素によって切断されます。 ヌクレアーゼ 5.8S rRNA、18S rRNA、28S rRNA の形成を伴います。
原核生物では、リボソーム RNA 分子は完全に異なる特性 (5S-、16S-、23S-rRNA) を持ち、これが発明および医学における多くの抗生物質の使用の基礎となっています。
転移RNA前駆体の処理
1. 脱アミノ化、メチル化、還元による分子内のヌクレオチドの修飾。
たとえば、プソイドウリジンとジヒドロウリジンの形成です。

修飾ウリジルヌクレオチドの構造
2. アンチコドンループの形成はスプライシングによって起こります
導入
タンパク質の生合成は、転写、プロセシング、翻訳の段階に分けることができます。 転写中に、DNA 分子内で暗号化された遺伝情報が読み取られ、この情報が mRNA 分子に書き込まれます。 一連の連続した処理段階で、後続の段階で不要な一部の断片が mRNA から除去され、ヌクレオチド配列が編集されます。 コードが核からリボソームに輸送された後、成長するポリペプチド鎖に個々のアミノ酸残基が結合することによって、実際のタンパク質分子の合成が行われます。
処理
転写と翻訳の間に、mRNA 分子は一連の連続的な変化を起こし、ポリペプチド鎖の合成のための機能マトリックスの成熟を確実にします。 真核細胞におけるプロセシングの出現により、遺伝子エキソンを組み合わせて、単一の DNA ヌクレオチド配列によってコードされるより多様なタンパク質を取得することが可能になりました。
キャッピング
キャップの化学構造
キャッピングが発生すると、転写物の 5" 末端に 5"-5" の異常な位置で接続する三リン酸架橋を介して 7-メチルグアノシンが付加され、最初の 2 つのヌクレオチドがリボースメチル化されます。キャッピング プロセスは、転写物の 5" 末端に追加されます。プレmRNA分子の転写の終わり。
キャップグループの機能:
- 核からのmRNA輸出の調節。
- 転写産物の 5" 末端をエキソヌクレアーゼから保護します。
- 放送開始への参加
ポリアデニル化
ポリアデニル化には、特殊な酵素ポリ (A) ポリメラーゼによって実行される、転写物の 3 インチ末端への 100 ~ 200 個のアデニル酸残基の付加が含まれます。
スプライシング
ポリアデニル化の後、mRNA はイントロンの除去を受けます。 このプロセスはスプライセオソームによって触媒され、スプライシングと呼ばれます。
放送
完成したタンパク質分子はリボソームから切断され、細胞内の目的の場所に輸送されます。 一部のタンパク質は、活性状態を達成するために追加の翻訳後修飾を必要とします。
ウィキメディア財団。 2010年。
他の辞書で「処理 (生物学)」が何であるかを確認してください。
この用語には他の意味もあります。「処理 (生物学)」を参照してください。 支払いを行うために必要な情報の処理と保管を含む処理活動。 この用語は銀行業界でよく使用されます... ... ウィキペディア
レンチウイルスベースのベクターを使用したヘアピンを含む低分子 RNA の送達と哺乳類細胞における RNA 干渉のメカニズム RNA 干渉 (... Wikipedia
ステムループを備えたプレ mRNA。 塩基の窒素原子は青で強調表示され、分子のリン酸骨格の酸素原子は赤で強調表示されます。リボ核酸 (RNA) は核酸、残基を含むヌクレオチドのポリマーです ... ... Wikipedia
分子生物学のセントラルドグマは、自然界で観察される遺伝情報の実装に関する規則を一般化します。つまり、情報は核酸からタンパク質に伝達されますが、その逆方向には伝達されません。 このルールはフランシスによって策定されました... ... ウィキペディア
ステムループを備えたプレ mRNA。 塩基の窒素原子は青で強調表示され、分子のリン酸骨格の酸素原子は赤で強調表示されます。リボ核酸 (RNA) は核酸であり、オルトリン酸残基を含むヌクレオチドのポリマーです ... Wikipedia
ステムループを備えたプレ mRNA。 塩基の窒素原子は青で強調表示され、分子のリン酸骨格の酸素原子は赤で強調表示されます。リボ核酸 (RNA) は 3 つの主要な高分子の 1 つです (他の 2 つは ... ウィキペディア
分子生物学のセントラルドグマは、自然界で観察される遺伝情報の実装に関する規則を一般化します。つまり、情報は核酸からタンパク質に伝達されますが、その逆方向には伝達されません。 このルールはフランシス・クリックによって策定されました... ... ウィキペディア
リボソームによるタンパク質合成のスキーム タンパク質生合成は、ポリペプチド鎖を合成する複雑な多段階プロセスです。
RNA合成(RNA転写)。
RNAの構造。
真核生物における遺伝物質の組織化。
遺伝情報の記録方法
遺伝物質の組織化。 ゲノムの機能部分。
遺伝子発現に関する一般的な情報。
1. 遺伝子発現に関する一般情報
ご存知のとおり、DNA には特定の遺伝情報が含まれています。
体のすべてのタンパク質とRNAの構造、および個体発生中およびさまざまな機能状態下でのさまざまな細胞におけるこの情報の実装順序について。
体のすべての体細胞は同じ 46 個の染色体セットを持っているため、細胞間の大きな違いにもかかわらず、すべての体細胞の DNA には同じ遺伝情報が含まれています。 (一部の例外はリンパ球であり、その形成中に免疫グロブリン遺伝子の再構成が起こります。)
DNA 複製中、遺伝情報は全体として再生され、娘細胞に受け継がれます。 しかし、さらに、この情報は細胞内で発現(実装)され、その生命活動のすべての症状を決定します。 ただし、核内の遺伝情報のすべてが発現されるわけではなく、その一部のみが発現されます。
特定のタンパク質の構造に関する情報の表現には、主に 2 つの段階があります。
a) それらの最初のものは転写です。細胞核内で、特別なメッセンジャーであるメッセンジャー RNA (mRNA) の対応する遺伝子 (染色体の 1 つに局在) が形成されます。
このプロセスの意味は、タンパク質の構造に関する情報を、巨大な固定担体 (染色体の一部としての DNA) から小さな可動担体である mRNA に書き換えることです。 数千のファイルが含まれるコンピュータのハード ドライブからそのうちの 1 つをフロッピー ディスクにコピーする場合も、状況はほぼ同じです。 したがって、遺伝子自体が互いに異なるのと同様に、異なる遺伝子から読み取られる mRNA も互いに異なるはずです。 もう 1 つの重要な状況: 遺伝子転写の直接産物は、より正確には mRNA の前駆体 (プレ mRNA) と呼ばれます。 実際のところ、新しく形成された mRNA はすぐに (核内で) 成熟またはプロセシングを受けます。 同時に大幅な変更も加えられています。 そしてその後初めて、成熟したmRNAが核から細胞質に入ります。
b) 遺伝子発現の主要な段階の 2 番目は翻訳です。mRNA によって指示されたプログラムに従って、リボソーム上でタンパク質が合成されます。 このプログラムの本質は、構築されるペプチド鎖にアミノ酸が含まれる順序を決定することです。 さらに、このプロセスには遊離アミノ酸ではなく活性化アミノ酸が含まれます。それらのそれぞれはいわゆるアミノ酸と関連付けられています。 トランスファー RNA (tRNA)、すなわち、アミノアシル tRNA (aa-tRNA) の形です。 20 個のアミノ酸のそれぞれに、tRNA の特定の形式があり、多くの場合、1 つではなく、複数の形式があります。
リボソームは翻訳において分子機械の役割を果たし、参加者の正しい相互作用を保証します。 リボソームには 4 つの分子が含まれています。 リボソーム RNA (rRNA) - 4 種類の rRNA のそれぞれ 1 分子。 リボソームタンパク質と結合することにより、それらはリボソームの 2 つのサブユニットを形成し、その中で構造的機能と、場合によっては触媒機能を実行します。 したがって、mRNA、tRNA、rRNA という 3 つのクラスの RNA が翻訳に関与します。
2. 遺伝物質の構成。 ゲノムの機能部分
遺伝子とその構造
タンパク質と RNA の構造に関する実際の情報は、遺伝子とシストロンと呼ばれる DNA のセクションに記録されています。
遺伝子は、1 つのタンパク質をコードする DNA のセクションです。
シストロン 1 つのポリペプチド鎖をコードする DNA の同じセクション。
動物と人間では、シストロンは異なる染色体上に存在することが多く、通常は遺伝子とも呼ばれます。 染色体には、体内のすべてのタンパク質の遺伝子に加えて、RNA 遺伝子 (4 種類のリボソーム RNA と数十の転移 RNA) も含まれています。
生物の遺伝情報を決定する遺伝子のセット全体を「遺伝子セット」といいます。 ゲノム.
ほぼすべての真核生物の遺伝子(原核生物の遺伝子とは異なります)には、コード領域だけが含まれるわけではないという特徴があります。 エクソン、ただしコーディング以外でも - イントロン。 エクソンとイントロンは互いに散在しており、これにより遺伝子に「壊れた」構造が与えられます。
遺伝子内のイントロンの数は 2 から数十まで変化します。 ミオシン遺伝子にはそのうちの約 50 個があり、場合によってはイントロンが遺伝子全長の最大 90% を占めることもあります。
DNAの他の部分
遺伝子の間には非コード配列もあります - スペーサー。 共通の名前にもかかわらず、機能的な役割はまったく異なる場合があります。
a) 多くのスペーサー領域が明らかに構造的な役割を果たしています。
ヌクレオソーム鎖を高次のクロマチン構造に正しく配置することに参加します。
染色体の中心小体装置などへの付着において。
b) DNA の他の非コード領域は、特定のタンパク質の特異的結合遺伝子座として機能します。
DNA上で機能する酵素
調節機能を果たすタンパク質。
この場合、RNAポリメラーゼ(DNA上でRNAを合成する酵素)の結合部位を次のように呼びます。 プロモーター。 それらは、遺伝子 (または遺伝子のグループ) の先頭に密接に隣接しているか、他の機能的遺伝子座によって遺伝子から分離されています。
c) 真核生物(ヒトを含む)では、遺伝子の「読み取り」の制御はリプレッサータンパク質だけでなく、いわゆるアクチベータータンパク質によっても行われます。 転写因子。
後者には、RNA ポリメラーゼをプロモーターに結合するために必要なすでに述べた一般的な転写因子が含まれます。 これらの因子はすべての細胞に存在し、機能する遺伝子を「読み取る」ために必要です。
他の転写因子は特定の遺伝子の活性のみを増加させ、そのような因子に結合する DNA 遺伝子座は、 エンハンサー.
d) 最後に、DNA には終結シグナルとして機能する短い遺伝子座が含まれる場合があります ( 終了) DNA 転写。
遺伝子が呼び出された後に位置する終結領域 ターミネーター.
3. 遺伝情報の記録方法
DNA鎖の機能的役割
遺伝子領域の 2 本の DNA 鎖は、その機能的役割が根本的に異なります。そのうちの 1 つは、 コーディングまたは セマンティック、 2番 - マトリックス.
これは、遺伝子を「読み取る」プロセス(転写、または mRNA 合成前)において、1 つの DNA 鎖、つまり鋳型だけが鋳型として機能することを意味します。 このプロセスの生成物であるプレ mRNA は、ヌクレオチド配列が DNA のコード鎖と同一です (チミン塩基がウラシル塩基に置換されています)。
したがって、DNA テンプレートの助けを借りて、DNA コード鎖の遺伝情報が転写中に RNA 構造内で再生されることがわかります。
図面では、コード鎖が上になるように遺伝子を描くのが通例です。 次に、DNA を描画するための一般的な規則に従って、コード鎖の 5 インチ末端が左側に配置される必要があります。
符号化回路の情報は5´→3´の方向に書き込まれます。 したがって、プロモーターは遺伝子のコード鎖の 5" 末端に位置します。そして、この同じ末端が遺伝子全体の 5" 末端であると考えられます (ただし、ここでは鋳型鎖には 3' 末端があります)。
遺伝暗号の基本的な性質
DNAコード鎖の情報の単位は、 三つ子- 3 つのヌクレオチドの配列。
DNA に含まれる 4 種類のヌクレオチドは、64 種類のトリプレットを形成できます。 これらのうち、61 個のトリプレットは意味論的です。つまり、それらは 20 個のアミノ酸のいずれかをコードしており、3 個のトリプレットは「意味がありません」です。
ご覧のとおり、平均して、アミノ酸ごとに意味上の三つ組がいくつかあります (実際には 1 から 6 まで)。 このため、遺伝暗号は次のように呼ばれます。 退化する。 これがなければ、非常に高い頻度でランダムな点突然変異(DNA 内の一部のヌクレオチドを他のヌクレオチドに置き換えること)が発生し、「意味のない」トリプレットが出現することになります。
同時にコード 特定の: センストリプレットのそれぞれは 1 つのアミノ酸のみに対応します。
タンパク質自体に関する情報は、完全な遺伝子 (イントロンを除く) では、トリプレットの直線配列が、このタンパク質の一次構造 (ペプチドのアミノからカルボキシル末端の方向) で同様のアミノ酸の直線配列をコードしているということです。鎖)。
タンパク質の一次構造がタンパク質分子の空間配置、およびその物理化学的および生物学的特性を決定するため、これで十分であることがわかります。
遺伝子のエクソンのトリプレットの配列とペプチド鎖のアミノ酸の間の線形対応は、次のように表されます。 共線性遺伝コード。
つまり、遺伝コードは三つ子です。 特定的、縮退的、共線的、連続的。 通常、このリストに追加されるのは 多用途性: すべての種類の生物において、どのトリプレットの意味も同じです。
遺伝コード
私たちがコードについて話すとき、これまでは DNA の意味論的な鎖を指してきました。 しかし、チミン (T) のウラシル (U) への置換を考慮すると、プレ mRNA のヌクレオチドの配列も同じです。
DNAのトリプレットに対応するmRNAのトリプレットは、 コドン。 実際、それらは直接的なものです:
リボソーム上で合成されるペプチド鎖にアミノ酸が含まれる順序が決まります。
1 つのアミノ酸のコドンは、最後の (3 番目) ヌクレオチドのみが異なります。
構造が似ているアミノ酸では、コドンも互いに似ています。つまり、それらは 2 つのヌクレオチド、または 1 つのヌクレオチド (ただし中心のヌクレオチド) に一致します。
4. 真核生物における遺伝物質の構成
多数のタンパク質とRNAの遺伝子
多くの真核生物の遺伝子の特徴の 1 つは、その構成中に非コード領域、つまりイントロンが存在することです。
もう 1 つの特徴は、固有の遺伝子 (半数体ゲノム内で単一のコピー数で表現される) に加えて、繰り返し反復される遺伝子が存在することです。
これら 2 つの特徴を説明するために、いくつかの特定の遺伝子を見てみましょう。
ヒストン遺伝子
ヒストン- クロマチンのヌクレオソーム構造の形成に関与する塩基性(酸塩基特性の観点から)タンパク質。 これらの 5 種類のタンパク質 (HI、H2A、H2B、H3、および H4) はそれぞれ、対応する遺伝子によってコードされています。
リボソームRNA遺伝子
リボソームには 4 種類の rRNA が含まれています。 これらの RNA は沈降定数が異なります。
遺伝子の機能は多くのタンパク質の影響を受けます。
一般的な転写因子
一般転写因子は、RNAポリメラーゼがプロモーターに結合するために必要な転写因子であり、それ自身もプロモーターと相互作用します。
転写因子としてのp53タンパク質
すでに発見されている多数の転写因子の中で、おそらく p53 タンパク質が最もよく知られています。 これは、それが非常に重要な細胞プロセスを制御しており、そのため、多数のさまざまな調節連鎖に関与しているという事実によって説明されます。
機能的な役割。
p53 タンパク質 (またはその遺伝子) は、細胞構造に対するさまざまな損傷に応答して活性化されます。
修復されていない切断およびその他の DNA 損傷
有糸分裂における染色体分離の違反、
微小管の破壊など
その結果、p53 タンパク質の仲介を通じて、細胞はその構造の損傷に反応します。
それは有糸分裂周期のいずれかの段階に留まり、これらの損傷を修復します。
あるいは(修正が不可能な場合)分裂が完全に停止し、細胞の老化プロセスに入ります。
あるいは、(損傷した細胞がその環境に対して潜在的に危険である場合)、アポトーシス、つまり簡単に言うと自殺を実行します。
特に、腫瘍の形質転換が起こった細胞はアポトーシスを起こします。 この点に関して、血管新生が同時に阻害される理由は明らかです。これは腫瘍の増殖を制限するもう 1 つの方法です。
したがって、p53 タンパク質は最も重要な腫瘍抑制因子の 1 つです。 進行中の腫瘍の大部分では、p53 タンパク質の機能が何らかの形で損なわれています。
5. RNAの構造
すべての転写因子は、転写自体と同様に、染色体の特定の部分で必要な速度で RNA を形成するという 1 つのことだけを保証するように設計されています。
RNAの構造の概略図
DNA と同様に、RNA は同じ構成原理を持つ直鎖状 (つまり、分岐していない) ポリヌクレオチドです。
4 種類のヌクレオチドで構成され、それぞれに窒素塩基、ペントース、リン酸残基が含まれます。
ヌクレオチドは、5',3'-ホスホジエステル結合を使用して鎖に結合されます。
ポリヌクレオチド鎖は極性です。つまり、識別可能な 5 インチ末端と 3 インチ末端を持っています。
しかし、DNAとは異なる点もあります。 主なものは、RNA 分子 (一部のウイルスの RNA を除く) は二本鎖ではなく一本鎖であるということです。 その理由は、一次構造の以下の3つの特徴にあります。
a) まず、RNA のペントースはデオキシリボースではなく、追加の水酸基を含むリボースです。 後者は二本鎖構造のコンパクト性を低下させます。
b) 第二に、4 つの主要な窒素含有塩基のうち、チミンの代わりにウラシルがあり、5 位にメチル基がない点のみがチミンと異なります。
6. RNA合成(DNA転写)
転写の一般的な特徴
DNA 複製とは異なり、DNA 転写は、分裂細胞と非分裂細胞の両方のほぼすべての有核細胞で発生します。
さらに、分裂細胞では、(真核生物の)複製期間と分裂そのものの期間を除いて、有糸分裂周期のどの時点でも起こります。
さらに、DNA の任意のセクションの転写は、サイクルのほぼすべての瞬間だけでなく、任意の回数、繰り返し発生する可能性があります。 一方、細胞内で転写される一連の領域は、特定の要因の影響を受けて変化することがよくあります。
このプロセスの酵素的サポートは、RNA ポリメラーゼによって行われます。 真核生物には 3 種類のこの酵素があります。
RNA ポリメラーゼ I - pre-rRNA の合成用。
RNA ポリメラーゼ II - プレ mRNA および
RNA ポリメラーゼ III - プレ tRNA 合成用
この酵素は DNA に沿って移動し、DNA 鋳型鎖のヌクレオチドに相補的なリボヌクレオチドの成長鎖への交互の組み込みを触媒します。
DNA 合成とのもう 1 つの類似点は、構築される鎖の成長方向 (5'→3') です。 これは、この鎖の次のヌクレオチドが 3 インチ末端に追加されることを意味します。
すべてのテンプレート合成と同様、構築される鎖は DNA テンプレート鎖と逆平行になります。 その結果、後者は酵素によって 3'→5' 方向に転写されます。
しかし、DNA 合成とは根本的に異なる点もあります。
a) プロセスの非対称性: ご存知のとおり、鋳型として使用される DNA 鎖は 1 つだけです。 酵素システムがどのようにして正しい鎖を選択するのかは完全には明らかではありません。 どうやら、ここで重要な役割を果たしているのは、システムによって認識される、鎖の 1 つ上のいくつかのヌクレオチド配列です。
b) 保存的プロセス: DNA 分子は RNA 合成が完了すると元の状態に戻ります。 DNA合成中、分子は半分更新されるため、複製は半保存的になります。
c) 最後に、RNA 合成の開始にはプライマーは必要ありませんが、DNA 複製には RNA プライマーが必要です。
転写メカニズム
転写の開始
転写の最初の、そしておそらく最も重要な段階は次のとおりです。 その開始: RNA ポリメラーゼのプロモーターへの結合と最初のヌクレオチド間結合の形成。
RNA ポリメラーゼの結合についてはすでに何度か説明したので、ここでは要点だけを思い出します (いくつかの情報を追加して)。
真核生物では複合体を形成するには、一般的な転写因子のタンパク質全体がプロモーターに事前に結合することが常に必要です。 RNA ポリメラーゼは、プロモーターに結合することにより、DNA の局所的な変性、つまり DNA の約 1.5 ターンにわたる DNA 鎖の分離を引き起こします。 彼らが言うように、転写の「目」が形成されます。 このおかげで、「目」領域の DNA テンプレート鎖のヌクレオチドが rNTP (リボヌクレオシド三リン酸) とペアリングできるようになります。
構築中の RNA 鎖に最初に含まれるのは常にプリン ヌクレオチド (ATP または GTP) であり、その 3 つのリン酸残基はすべて保持されます。
次に、最初の 5",3" リン酸結合が 2 番目のヌクレオチドと形成されます。
転写伸長
開始後の次の段階は伸長です。成長するプレ RNA 鎖が最終サイズまで徐々に長くなります。
これは、RNA ポリメラーゼが DNA に沿って移動するときに発生します。 それに応じて、転写の「目」、つまり局所的なDNAがほどける領域も動きます。 DNA の転写部分では、RNA ポリメラーゼの出発直後に二本鎖らせん構造が復元されます。
酵素の移動と RNA 合成のおおよその速度は 1 秒あたり 30 ヌクレオチドです。
転写の終了
最後の段階は終了、つまり転写の終了です。
このシグナルは、遺伝子の末端にある特別な GC リッチ領域です。 GC ペア間の相互作用力は非常に強いため、DNA 内のそのような領域の局所的な変性はより困難になります。 これは RNA ポリメラーゼの進行を遅らせ、転写を停止する信号として機能します。
しかし、プロセスが終了する前であっても、新しく合成された RNA の末端には GC に富む領域が出現します。 ヌクレオチド間の相互作用により、「ヘアピン」を形成します。
つまり、DNA テンプレート鎖のヌクレオチドとの相互作用が「ヘアピン内」相互作用に置き換えられます。 これにより、RNA が DNA から剥がれやすくなります。
7. RNAの成熟(処理)
ほぼすべての RNA 成熟プロセスは 3 つのタイプに分類できます。
一部を削除
他の人に加わったり、
同じヌクレオチドまたは 3 番目のヌクレオチドの修飾。
「余分な」シーケンスの削除
概要
「余分な」ヌクレオチドの除去は、特別なヌクレアーゼによって実行されます。 エキソヌクレアーゼは、鎖の特定の末端 (3' または 5') から一度に 1 ヌクレオチドを順番に切断します。 そして、エンドヌクレアーゼは中間セクションのどこかで鎖を切断し、その断片化を引き起こします。
スプライシングの仕組み
検討中のメカニズムの重要なポイントの 1 つは、プレ RNA 鎖の切断の精度を確保することです。ヌクレオチドが 1 つでもあると「フレーム シフト」が発生し、すべての mRNA コドンまたは tRNA アンチコドンの意味が変化します。 。
精度は次の 2 つの状況によって達成されます。
まず、各イントロンの最初と最後には特定のヌクレオチド配列があります。たとえば、イントロンは常に G-U で始まり、ダブレット A-G で終わります。
次に、これらの配列を認識するために、特別な RNA が使用されます。 核内低分子RNA(snRNA)。 後者はスプライシングを触媒する酵素と関連しています。 このようなリボ核タンパク質複合体はスプリオソームと呼ばれます。
スプライシングは、イントロンの最初と最後にある 2 つの snRNA の相互作用から始まります。 これにより、エンドヌクレアーゼの「方向」が決まります。エンドヌクレアーゼは二本鎖領域と一本鎖領域の境界で作用します。
最初の pre-RNA 切断は、イントロンの 5' 末端の領域で発生します。これは、左側の snRNA の左端の位置です。 この場合、イントロンの 5 インチ末端が同じイントロンの中央部分にあるヌクレオチドの 1 つに結合し、これにより環構造が形成されます。
ヌクレオチドの付加および修飾
したがって、プレRNAの成熟の過程で、後者はそのヌクレオチドのかなりの部分を失います。 しかし、個々のヌクレオチドの非転写的な付加も起こります。
プレ mRNA の場合、「キャップ」の構成要素である 7-メチルグアニル ヌクレオチドが 5 インチ末端に結合します (ピロリン酸結合を使用しますが、これはポリヌクレオチドでは一般的ではありません)。そして 3 インチ末端には、約 200 ヌクレオチドのポリ (A) フラグメントがヌクレオチドごとに拡張されます。 この目的のために、特別な酵素が使用されます。 特に、ポリアデニレートポリメラーゼのポリ(A)フラグメントの形成に使用されます。
pre-tRNA の場合、3 つのヌクレオチド (C、C、A) が 3" 末端から順番に追加され、アクセプター分岐を形成します。